Polimorfismo R577X de ACTN3 y Respuesta Neuromuscular al Entrenamiento con Sobrecarga
Martín F Bottaro1, Paulo Gentil1, Rinaldo W. Pereira2, Tailce K. Leite21 College of Physical Education, University of Brasilia, Brasil.
2 Genomic Sciences and Biotechnology, Catholic University of Brasilia, Brazil.
Resumen
Al polimorfismo R577X del gen ACTN3 se lo ha asociado con la fuerza muscular, la hipertrofia y el estado atlético. Se cree que el alelo X, al que se lo asocia con la ausencia de la proteína del ACTN3, desmejora el rendimiento en contracciones musculares de alta fuerza/velocidad. El propósito de este estudio fue investigar la asociación entre el polimorfismo R577X con la respuesta muscular al entrenamiento con sobrecarga en hombres jóvenes. Ciento cuarenta y un hombres realizaron dos sesiones de entrenamiento con sobrecarga por semana durante 11 semanas. A los participantes se les evaluó la fuerza en 1 RM en el ejercicio de press de banca, el torque pico de los extensores de la rodilla y el grosor muscular de los extensores de la rodilla antes y después del período de entrenamiento. La genotipificación se llevó a cabo utilizando la enzima de restricción DdeI. La distribución de los genotipos fue del 34.4% para el genotipo RR, el 47% para el RX y el 18.6% para el XX. De acuerdo con estos resultados, el polimorfismo R577X en el gen ACTN3 no está asociado con la fuerza muscular inicial ni con la respuesta de la fuerza muscular al entrenamiento con sobrecarga. No obstante, solo los portadores del alelo R mostraron incrementos en el grosor muscular en respuesta al entrenamiento.
Palabras Clave: fuerza muscular, hipertrofia muscular, torque pico, genotipo, alfa-actinina 3, extensor de la rodilla
INTRODUCCIÓN
La fuerza muscular ejerce una influencia importante sobre las capacidades funcionales y se la ha asociado de manera positiva con el rendimiento deportivo (ACSM, 2009; Jung, 2003), la longevidad y la calidad de vida (Newman et al., 2006; Visser et al., 2005). Aunque se sabe que los estímulos del entorno son factores determinantes de la fuerza y el tamaño muscular, la herencia tiene una influencia significativa sobre estos fenotipos (Bray et al., 2009; Stewart y Rittweger, 2006; Thomis et al., 1997). Se estima que los factores genéticos pueden corresponder del 44 al 58% de las variaciones inter-individuo en la fuerza muscular (Beunen y Thomis, 2004; Tiainen et al., 2009) y la masa magra (Arden ay Spector, 1997), y esta influencia es más evidente en las personas jóvenes que en las mayores (Prior et al., 2007; Stewart y Rittweger, 2006). En este aspecto, uno de los genes que puede influenciar estos fenotipos es el gen alfa-actinina 3 (ACTN3) (Clarkson et al., 2005; Niemi y Majamaa, 2005; Yang et al., 2003).
La ACTN3 es una proteína específica de las fibras tipo 2 (Beggs et al., 1992; North et al., 1999), ubicada en los discos Z, y tiene funciones importantes en el metabolismo y la estructura muscular (Beggs et al., 1992; MacArthur y North, 2004, 2007). El gen ACTN3 está ubicado en la banda cromosómica 11q13-q14 y fue clonado por primera vez por Beggs et al. (1992). Posteriormente, North et al. (1999) identificaron un polimorfismo funcional en el gen ACTN3 en los seres humanos. Esta mutación, conocida como R577X (rs1815739), es una transición de la citosina (alelo R) a la timina (alelo X) en la posición 1747 en el exon 16, dando como resultado un codón de terminación prematuro, lo que está asociado con la ausencia de la expresión de la ACTN3 en los portadores del genotipo XX (North et al., 1999).
El genotipo XX no provoca ningún fenotipo aparente ni cambios histológicos, lo que sugiere que la presencia de la proteína no es crítica (North y Beggs, 1996) y las variaciones en la función muscular se producen dentro del rango normal (MacArthur et al., 2008). Análisis realizados en ratas revelaron que la ausencia de la ACTN3 llevó a la pérdida de masa muscular, asociada con reducciones en el área de sección cruzada de los músculos compuestos principalmente por las fibras de tipo 2 (Chan et al., 2008; MacArthur et al., 2008). Además, las fibras de animales con el genotipo XX tienen una capacidad de trabajo menor y un mayor tiempo medio de relajación (Chan et al., 2008; MacArthur et al., 2008).
Estudios previos han demostrado que el genotipo XX tiene escasa representación en los atletas de fuerza y potencia en relación con los controles y los atletas de resistencia (Druzhevskaya et al., 2008; Eynon et al., 2009; Niemi y Majamaa, 2005; Papadimitriou et al., 2008; Roth et al., 2008; Yang et al., 2003; Yang et al., 2007). Sobre la base de la función fisiológica de la ACTN3 y la relación entre las formas polimórficas del gen ACTN3 y el rendimiento del ejercicio, se espera que el polimorfismo R577X esté asociado con los valores iniciales del tamaño muscular, la fuerza y la potencia; no obstante, los resultados de los estudios de diseño transversal son controversiales (Clarkson et al., 2005; Delmonico et al., 2007; Delmonico et al., 2008; McCauley et al., 2009; Moran et al., 2007; Norman et al., 2009; San Juan et al., 2006; Walsh et al., 2008). Estudios realizados en personas mayores (Delmonico et al., 2008; San Juan et al., 2006; Walsh et al., 2008) y más jóvenes (McCauley et al., 2009; Moran et al., 2007; Norman et al., 2009) revelaron que no hubo diferencias en la fuerza y la potencia muscular entre los diferentes genotipos R577X. Sin embargo, algunos análisis por separado realizados en mujeres revelaron que los portadores del genotipo XX tienen menos fuerza y potencia muscular (Clarkson et al., 2005; Delmonico et al., 2008; Walsh et al., 2008), mientras que otros reportaron que el grupo del genotipo XX tiene una mayor potencia muscular que los portadores del alelo R (Delmonico et al., 2007).
Si bien se sabe que los factores genéticos pueden influenciar la respuesta de la fuerza muscular al entrenamiento con sobrecarga, la determinación de los genes específicos asociados con esta respuesta aún está en sus inicios. Con respecto a la ACTN3, Delmonico et al. (2007) reportaron que las mujeres mayores, pero no los hombres, con genotipo RR tuvieron mayores incrementos en la potencia muscular después de 10 semanas de entrenamiento unilateral con sobrecarga de los extensores de la rodilla. No se observaron diferencias en las ganancias de 1RM entre los genotipos en ninguno de los géneros. El único estudio realizado con sujetos jóvenes fue publicado por Clarkson et al. (2005). Los autores estudiaron las asociaciones entre el genotipo ACTN3 y la fuerza del flexor del codo en hombres y mujeres jóvenes que participaron de 12 semanas de entrenamiento con sobrecarga. Se reportó que las mujeres portadoras del alelo X tuvieron mayores ganancias en 1RM, en comparación con los homocigotos RR, aunque los resultados en los hombres no fueron significativos (Clarkson et al., 2005). Por lo tanto, no se sabe si el alelo R favorece las adaptaciones musculares debido a su función fisiológica o si los portadores del alelo X se ven favorecidos por el “principio del valor inicial”, lo que determina que los participantes con valores iniciales más bajos muestran una mayor respuesta al entrenamiento, según sugirieron Clarkson et al. (2005). Debido a la controversia en la literatura, como también a la escasez de estudios experimentales que investigan los efectos del R577X en la fuerza e hipertrofia muscular en respuesta al entrenamiento, el propósito del presente estudio fue investigar la influencia del polimorfismo en el gen ACTN3 sobre la respuesta muscular al entrenamiento con sobrecarga en hombres jóvenes.
MÉTODOS
Visión General del Estudio
El estudio investigó la asociación del genotipo R577X de ACTN3 con la respuesta neuromuscular al entrenamiento con sobrecarga en 141 hombres jóvenes. Antes y 5-7 días después de las 11 semanas de entrenamiento, se evaluó la fuerza en 1RM en el ejercicio de press de banca y el torque pico concéntrico de los extensores de la rodilla de todos los voluntarios. Las sesiones de entrenamiento con sobrecarga incluyeron dos series de 8-12 repeticiones de cinco ejercicios y fueron diseñadas para ejercitar los principales grupos musculares. Todas las sesiones de prueba y entrenamiento se llevaron a cabo en el mismo momento del día. Para el análisis del genotipo, se recolectaron 4 mL de sangre de la vena antecubital después del último test de RM en press de banca. La genotipificación se llevó a cabo utilizando un procedimiento que incluyó la enzima de restricción DdeI.
Participantes
Doscientos diez hombres universitarios participaron de manera voluntaria en el estudio. A los voluntarios se los seleccionó de manera aleatoria entre aquellos que respondieron a la distribución de volantes realizada en el campus de la universidad, y por el boca a boca. Los criterios para el ingreso al estudio incluyeron tener al menos 18 años de edad, no contar con experiencia previa en entrenamiento con sobrecarga y no tener problemas clínicos que pudieran agravarse por los procedimientos del estudio. Para ser incluidos en el estudio, los individuos debieron asistir al menos al 90% de las sesiones de entrenamiento. A los voluntarios se los orientó a no cambiar sus hábitos nutricionales durante el período de estudio, y en caso de detectarse cualquier cambio relevante (i.e. convertirse en vegetariano, restricción de calorías, tomar suplementos nutricionales o ácidos ergogénicos, etc.) los datos de los participantes se excluían del análisis. Al final del estudio, 141 sujetos reunieron los criterios para ingresar al estudio (22.0 ± 2.7 años; 1.75 ± 0.07 m; 72.0 ± 13.9 kg). La mayor parte de las exclusiones se debieron al rendimiento en el entrenamiento con sobrecarga, además del protocolo del estudio, los cambios en los hábitos nutricionales y la baja concurrencia al entrenamiento. Los individuos excluidos eran similares a los otros con respecto a la distribución de los genotipos y las características físicas. El estudio se realizó según los estándares éticos del International Journal of Sports Medicine (Harriss y Atkinson, 2009). A todos los participantes se les informó sobre los procedimientos, requerimientos, beneficios y riesgos de la investigación antes de que dieran el consentimiento informado por escrito. El Comité Institucional de Ética de la Investigación aprobó el estudio.
Procedimientos
Test de 1 Repetición Máxima
La semana previa al experimento y 5-7 días después de la última sesión de entrenamiento, se determinó la carga de 1RM en el ejercicio de press de banca para cada sujeto. El protocolo de prueba recomendado por Kraemer y Fry (1995) se adoptó de la siguiente manera: (1) Una entrada en calor consistente en 5-10 repeticiones al 40-60% de 1RM estimada, (2) Un minuto para descansar con un leve estiramiento, seguido de 3-5 repeticiones al 60-80% de 1RM estimada, (3) Tres a cinco intentos de alcanzar la 1RM con intervalos de descanso de 5 minutos entre cada nuevo levantamiento. Se registró el peso máximo levantado de manera exitosa. Durante las pruebas se proporcionó estímulo verbal para obtener el mejor rendimiento. Las pruebas iniciales se repitieron en todos los sujetos, y el coeficiente de correlación intra-clase (ICC) fue de 0.98.
Medición del Torque Pico Isocinético Concéntrico (PT)
El PT isocinético concéntrico del extensor de la rodilla se midió en un Dinamómetro Isocinético Biodex System 3 (Biodex Medical, Inc., Shirley, NY). El dinamómetro se calibró según las especificaciones del fabricante antes de cada sesión de prueba. Los participantes se sentaron erguidos con el eje de rotación del brazo del dinamómetro orientado con el cóndilo femoral lateral de la rodilla derecha. Se utilizaron cinturones para asegurar el muslo, la pelvis y el tronco a la silla del dinamómetro a fin de prevenir el movimiento adicional del cuerpo. Se registró la posición de la silla y del dinamómetro para asegurar la misma ubicación para todas las pruebas. La corrección de la gravedad se obtuvo mediante la medición del torque ejercido sobre el adaptador de resistencia del dinamómetro con la rodilla relajada en una extensión casi total. Las pruebas incluían dos series de cinco repeticiones a 60º/s. A los sujetos se les ordenó que extendieran y flexionaran por completo la rodilla y trabajaran al máximo durante cada serie. Se proporcionó estímulo verbal durante toda la sesión de prueba. Después de cada serie, fue necesario que los sujetos tomaran 60 s de descanso antes de comenzar la serie siguiente. La correa de la rodilla se retiró en cada periodo de descanso a fin de asegurar un flujo sanguíneo sin restricción hacia el miembro inferior. El mismo investigador administró los procedimientos a todos los participantes. El ICC de la prueba y el retest de los valores iniciales del PT del extensor de la rodilla y el error estándar de la media (SEM) fueron de 0.98 y 2.3 % respectivamente.
Grosor Muscular (MT)
A los participantes se los evaluó antes y después del período de entrenamiento de 11 semanas para determinar el MT de los extensores de la rodilla de la extremidad derecha. Todas las pruebas se llevaron a cabo en el mismo momento del día, y a los participantes se les ordenó que se hidrataran normalmente 24 horas antes de las pruebas. Las mediciones se llevaron a cabo 3-5 días después de la última sesión de entrenamiento a fin de prevenir cualquier hinchazón por participar de la medición de MT (Chilibeck et al., 2004). Durante este período, a los participantes se les ordenó que no participaran de ninguna otra sesión de ejercitación o actividad intensa. El MT se midió utilizando un ultrasonido Modo B (Philips-VMI, Ultra Vision Flip, modelo BF). En el lugar de la medición se aplicó un gel de transmisión soluble en agua y se colocó una sonda de ultrasonido de 7.5 MHz perpendicular a la interfaz del tejido sin presionar la piel. Los puntos anatómicos se seleccionaron según Bemben (2002). Una vez que el técnico se sentía satisfecho con la calidad de la imagen producida, congelaba la misma en el monitor. Con la imagen congelada, un cursor podía medir el MT, que se tomaba como la distancia desde la interfaz tejido-músculo de la adiposidad subcutánea hasta la interfaz músculo-hueso (Abe et al., 2000). Un técnico calificado realizó todos los análisis (Sanada et al., 2006). El coeficiente de variación fue menor al 3.0%. El ICC de la prueba y retest de los valores iniciales fue de 0.99.
Intervención del Entrenamiento con Sobrecarga
Las sesiones del entrenamiento con sobrecarga consistieron de cinco ejercicios que incluían press de pierna, flexión de rodilla, press de banca, tirones en polea y abdominales. A fin de mejorar la validez ecológica y seguir las recomendaciones de la literatura (ACSM, 2009), los participantes realizaron dos series de 8-12 repeticiones. La intensidad del entrenamiento se estandarizó para todos los grupos, pues, inicialmente se les ordenó a los sujetos que entrenaran con ocho repeticiones máximas (8RM) y progresaran hasta mantener una carga de 8-12 RM. Todas las series se llevaron a cabo hasta el fallo concéntrico. Al progresar el entrenamiento, el técnico en ejercicios incrementaba la sobrecarga cuando un participante completaba 12 repeticiones durante al menos una de las dos series mientras que mantuviera la forma adecuada. Todos los participantes siguieron el mismo programa de entrenamiento con sobrecarga durante 11 semanas bajo supervisión. Los entrenadores expertos supervisaron detenidamente las sesiones de entrenamiento, pues la investigación previa ha demostrado mayores ganancias en el entrenamiento supervisado contra el no-supervisado (Gentil y Bottaro, 2010).
El entrenamiento se llevó a cabo dos días a la semana, con un mínimo de 48 horas entre las sesiones. Las series comenzaban cada tres minutos, seguidas de un intervalo de descanso de aproximadamente dos minutos. A los participantes se les indicó que llevaran registros de entrenamiento para cada día de trabajo. Después de cada sesión de ejercicios, un investigador/supervisor completaba y verificaba todos los registros de entrenamiento del estudio de 11 semanas.
Genotipificación
El ADN se extrajo utilizando un protocolo de remoción de sales. Se obtuvieron muestras de sangre de la vena antecubital de todos los voluntarios en tubos que contenían anticoagulante EDTA.
El locus genómico que contenía el lugar polimórfico se amplificó utilizando PCR. Los cebadores para PCR (5’-CCCACAACTTTAGGCTCCTG-3’ – directo y 5’-ATGTAGGGATTGGTGGAGCA-3’ – inverso) se diseñaron utilizando el programa Primer3Plus, disponible en Internet (http://www.bioinformatics.nl/cgi-bin/primer3plu s/primer3plus.cgi). Luego, al fragmento amplificado de 342pb se lo sometió a digestión por medio de DdeI (Promega, Madison, WI, EUA). Más tarde, a los productos digeridos se los sometió a una electroforesis en un 2% de gel de agarosa y se los visualizó en luz UV. El análisis visual del gel dio como consecuencia tres posibles resultados: RR (342pb), RX (342, 250, 92pb) o XX (250, 92pb). La precisión del ensayo de genotipificación se verificó utilizando muestras de control positivas y negativas previamente genotipificadas, utilizando el mismo método y siguiendo las recomendaciones previas (Chanock et al., 2007).
Análisis Estadísticos
La distribución de los genotipos R577X de ACTN3 se analizó por medio del análisis Ji Cuadrado para verificar la concordancia con el equilibrio de Hardy-Weinberg. Para evaluar diferencias entre los genotipos respecto de la edad, la altura, el peso, el valor inicial de la fuerza en 1RM en press de banca y el PT y MT de los extensores de la rodilla, se realizó un Análisis de Varianza (ANOVA). A fin de analizar la asociación entre los genotipos R577X de ACTN3 y las adaptaciones inducidas por el RT, se utilizó el análisis de varianza ANOVA para medidas repetidas (genotipo × tiempo), en el que los factores intra-sujeto fueron los valores pre- y post- de los fenotipos en estudio y los factores entre los sujetos fueron los genotipos (RR, RX y XX). Cuando fue necesario, las comparaciones múltiples con ajustes de intervalos de confianza mediante el procedimiento de Bonferroni se utilizaron como post hoc. Se calculó el cambio de porcentaje relativo para el MT del extensor de la rodilla, el PT y 1RM de press de banca utilizando la siguiente ecuación: [(Valores Post – Valores Pre)/Valores Pre x 100]. Los datos se consideraron significativos si p < 0.05 y los análisis estadísticos se llevaron a cabo utilizando el Programa Estadístico para Ciencias Sociales 16.0 (SPSS, Chicago, IL). Los datos están expresados en medias ± desviación estándar.
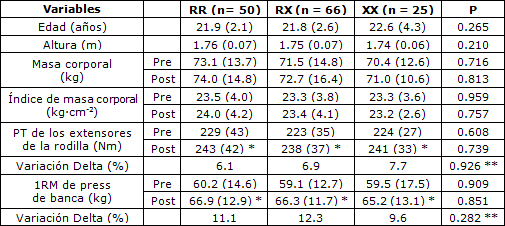
Tabla 1. Características de los participantes según los genotipos R577X de ACTN3. Los valores son medias (± desviación estándar). PT= torque pico; 1RM= una repetición máxima; *p < 0.05 entre pre y post-entrenamiento; **tiempo por interacción de genotipo.
RESULTADOS
En la Tabla 1 se presentan las características de los participantes según el genotipo R577X de ACTN3. La distribución de los genotipos fue en equilibrio de Hardy-Weinberg (χ2 = 0.726 p > 0.05). Según los resultados, el 35.5% de los voluntarios se clasificaron como RR, el 46.8% como RX y el 17.7% como XX. El cumplimiento general del programa de entrenamiento fue del 93% y no difirió entre los genotipos.
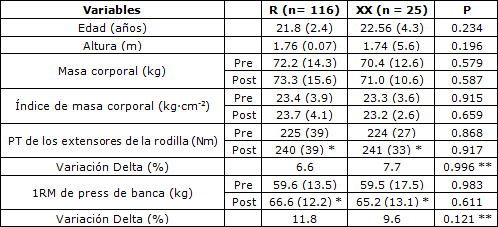
Tabla 2. Características de los participantes según los genotipos R577X de ACTN3. Los valores son medias (± desviación estándar). PT= torque pico; 1RM= una repetición máxima; *p < 0.05 entre pre y post-entrenamiento; **tiempo por interacción de genotipo.
No hubo diferencias entre los genotipos para los valores iniciales del PT de los extensores de la rodilla y la fuerza en 1RM en press de banca (p > 0.05). El ANOVA para el PT y 1RM no reveló ninguna interacción significativa de grupo por tiempo (p > 0.05). No obstante, se halló un efecto principal significativo para el tiempo para el PT y 1RM. Al analizarse el efecto principal de tiempo, todos los genotipos aumentaron de manera significativa el PT de los extensores de la rodilla y 1RM de press de banca (p < 0.05). Las comparaciones adicionales entre los portadores del alelo R (RR + RX) y el genotipo XX (Tabla 2) no revelaron ninguna diferencia entre los grupos para los valores iniciales y los cambios en el PT de los extensores de la rodilla y 1RM de press de banca (p > 0.05).
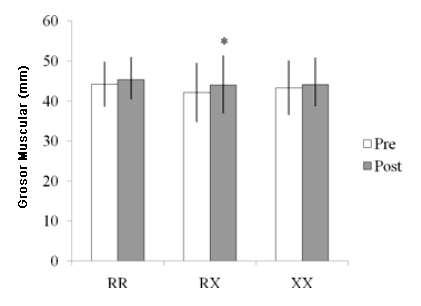
Figura 1. Medidas del grosor muscular de los extensores de la rodilla pre- y post-entrenamiento según el genotipo R577X. *Diferencia significativa entre pre- y post-entrenamiento (p < 0.05).
Se dispuso de las mediciones del MT de 40 participantes (edad 20.9 ± 2.6 años; altura 1.75 ± 0.08 m; peso 70.5 ± 14.9 kg; 1RM de press de banca 59.0 ± 10.1 kg; y PT de los extensores de la rodilla 221.18 ± 33.49 Nm). Treinta y uno fueron clasificados como R y nueve como XX. Los análisis por separado de estos participantes con respecto a las distribuciones del genotipo y los resultados de la prueba revelaron que sus características no diferían de la muestra completa. Además, la distribución del genotipo fue similar al grupo completo (30% para RR, 50% para RX y 20 para XX). El ANOVA para el MT no reveló ninguna interacción significativa de grupo por tiempo (p > 0.05). Sin embargo, se halló un efecto principal significativo para el tiempo para el MT. Cuando se analizó el efecto principal de tiempo, solo el genotipo RX aumentó de manera significativa el MT (p < 0.05; Figura 1). Al realizarse las comparaciones entre los portadores y los no-portadores del alelo R, solo los portadores del alelo R mostraron incrementos significativos en el MT de los extensores de la rodilla (p < 0.05; Figura 2). Un tamaño de muestra de 40 sujetos (30 para R, y 9 grupos de XX) proporcionaron > 0.80% de poder estadístico a un nivel α de 0.05 (2-colas) para las mediciones del MT.
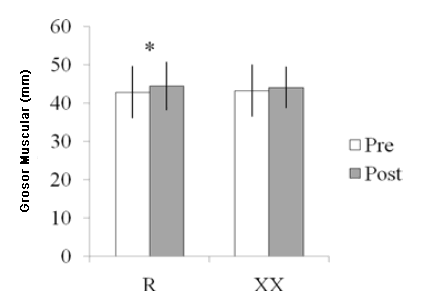
Figura 2. Medidas del grosor muscular de los extensores de la rodilla pre- y post-entrenamiento en los portadores del alelo R y el genotipo XX. *Diferencia significativa entre pre- y post-entrenamiento (p < 0.05).
DISCUSIÓN
La distribución de genotipos en el presente estudio fue similar a la reportada anteriormente en caucásicos (Clarkson et al., 2005; Delmonico et al., 2007; 2008; Druzhevskaya et al., 2008; Eynon et al., 2009; Moran et al., 2007; Niemi y Majamaa, 2005; Paparini et al., ; Walsh et al., 2008) e hispánicos (Clarkson et al., 2005), pero diferentes a la distribución reportada en asiáticos y afro-americanos, que presentan una frecuencia incrementada y disminuida del alelo X respectivamente (Clarkson et al., 2005; North y Beggs, 1996). En el presente estudio, se evaluó si el polimorfismo R577X estaba asociado a la fuerza muscular de los valores iniciales y el MT y su respuesta al entrenamiento con sobrecarga en un grupo de hombre jóvenes sin entrenamiento. La fuerza en 1RM en el ejercicio de press de banca, el PT de los extensores de la rodilla y el MT de los extensores de la rodilla se midieron antes y 11 semanas después de un protocolo de entrenamiento con sobrecarga.
Se halló que los valores iniciales de la fuerza muscular y el MT no fueron diferentes entre los genotipos de R577X, lo que concuerda con los análisis realizados en hombres mayores y jóvenes (Delmonico et al., 2007; 2008; McCauley et al., 2009; Norman et al., 2009; San Juan et al., 2006).
Los análisis que reportaron diferencias en la fuerza y la potencia muscular entre los genotipos de ACTN3 se llevaron a cabo con mujeres (Clarkson et al., 2005; Delmonico et al., 2007; Walsh et al., 2008), sugiriendo que los efectos del polimorfismo R577X puede ser dependiente del género. Una explicación posible para este efecto dependiente del género es que el efecto anabólico de las hormonas masculinas esteroides podría superponer los efectos de las ganancias de la fuerza y potencia muscular como resultado de la proteína ACTN3 en la estructura muscular (MacArthur y North, 2004).
Con respecto a la respuesta de la fuerza y la potencia muscular al entrenamiento con sobrecarga, Clarkson et al. (2005) estudiaron la asociación de los genotipos R577X con los fenotipos musculares en hombres y mujeres jóvenes, antes y después de 12 semanas de entrenamiento con sobrecarga. El entrenamiento se llevó a cabo dos días por semana y estuvo dirigido a los flexores y extensores del codo del lado no dominante. De acuerdo con los resultados, las mujeres que portan el alelo X mostraron incrementos mayores de 1RM que las que portan el genotipo XX. Sin embargo, los resultados en los hombres no alcanzaron una significancia. Más tarde, Delmonico et al. (2007) examinaron los efectos del polimorfismo R577X en los cambios de la potencia muscular después de 10 semanas de entrenamiento con sobrecarga en mujeres y hombres mayores. El entrenamiento se llevó a cabo tres veces por semana y estaba compuesto de ejercicios para los extensores y flexores de la rodilla. En contraposición a los resultados reportados por Clarkson et al. (2005), las mujeres con el genotipo RR tuvieron incrementos en la potencia muscular, pero los resultados en los hombres no alcanzaron una significancia. No hubo diferencias en el incremento de 1RM entre los genotipos para los hombres y las mujeres. La razón para las diferencias entre los resultados de Delmonico et al. (2007) y Clarkson et al. (Clarkson et al., 2005) puede estar relacionada con las características de los grupos, pues en ambos estudios, el grupo con valores iniciales más bajos mostró incrementos mayores, independientemente de los genotipos. Por lo tanto, los resultados pueden estar relacionados a una mayor posibilidad de mejoras en personas con valores iniciales más bajos y no a las variaciones en el genotipo ACTN3.
Es importante destacar que en los estudios de Clarkson et al. (2005) y Delmonico et al. (2007) no hubo diferencias para los cambios en el rendimiento entre los genotipos en los hombres. Por lo tanto, los presentes resultados confirman los estudios previos realizados con hombres (Clarkson et al., 2005; Delmonico et al., 2007). Aunque el presente estudio y otros estudios experimentales y de diseño transversal sugieren que el polimorfismo R577X no influencia los fenotipos musculares en los hombres, el hecho de que el genotipo XX tenga escasa representación en los atletas de fuerza y potencia (Druzhevskaya et al., 2008; Eynon et al., 2009; Niemi y Majamaa, 2005; Papadimitriou et al., 2008; Roth et al., 2008; Yang et al., 2003; 2007) es un enigma. Si el éxito de los atletas de deportes que requieren contracciones musculares de fuerza elevada y alta intensidad al parecer está relacionado con la presencia de la proteína ACTN3, es posible sugerir que el alelo R es favorable para generar fuerza y velocidad, y que este alelo puede estar asociado con una mejor respuesta al entrenamiento. No obstante, esto no se vio en el presente estudio.
Una explicación posible es que, en los participantes no atletas, la diferencia que deriva del polimorfismo R577X, cuando se lo analiza solo, es demasiado baja para ser relevante, en especial en la gente joven, según lo que sugieren McCauley et al. (2009). Los análisis previos han sugerido que el genotipo R577X es responsable del ~1-2% de las variaciones en la fuerza muscular (Clarkson et al., 2005; Walsh et al., 2008), aunque esta diferencia parece relativamente pequeña en las actividades de la vida diaria, puede ser la diferencia entre ganar o perder una competencia en un contexto de alta competición. Además, para un atleta de alto nivel, es necesario tener una combinación de genes favorables a fin de ser exitoso, y la ACTN3 puede ser uno de estos tantos genes. Por lo tanto, al analizar a los atletas, es probable que no solo se estén analizando los portadores del alelo R, sino, que se estén estudiando los portadores de una reserva genética favorable, entre ellos está el genotipo R577X. Por otro lado, es posible que este no sea el caso cuando se estudia una muestra de no-atletas, la que puede tener características genéticas heterogéneas, tales como que el alelo R no esté acompañado de otras variantes genéticas favorables.
Además, la intensidad y el volumen de las sesiones de entrenamiento realizadas en este y otros estudios (Clarkson et al., 2005; Delmonico et al., 2007) pueden haber sido demasiado bajos en comparación con el entrenamiento que realizan los atletas. A menudo los atletas entrenan muchas horas al día durante muchos años, mientras que los estudios disponibles no duran más de 12 semanas y no incluyen más de 3 horas de entrenamiento por semana. Este entrenamiento de bajo volumen y corta duración no pudo haber sido suficiente para inducir la manifestación de las diferencias funcionales entre los genotipos R577X.
Con respecto a los efectos de las variaciones del gen ACTN3 en la hipertrofia muscular en respuesta al entrenamiento, Clarkson et al. (Clarkson y Newham, 1995) no hallaron diferencias entre los genotipos R577X en la respuesta de la masa muscular a 12 semanas de entrenamiento con sobrecarga en hombres y mujeres jóvenes. En concordancia con los resultados de Clarkson y Newham (1995), tampoco se hallaron diferencias entre los genotipos en los presentes análisis. Sin embargo, es importante mencionar que los portadores del alelo R mostraron incrementos significativos en el MT en respuesta al entrenamiento.
El hecho de que la ausencia del alelo R afecta la hipertrofia muscular en respuesta al entrenamiento podría deberse a los efectos funcionales del ACTN3. Yang et al. (2003) inicialmente sugirieron que la proteína ACTN3 promueve la formación de las fibras de tipo II. Más tarde, Vincent et al (Vincent et al., 2007) demostraron que los portadores del genotipo XX tenían un porcentaje menor de las fibras tipo IIx en comparación con los portadores del alelo R, aunque el estudio de Norman et al. (Norman et al., 2009) no concuerda con esto. Si al genotipo XX, de hecho, se lo asocia con una proporción más baja del tipo de fibra rápida, es comprensible que este grupo muestre una limitación en la hipertrofia, pues estudios previos demostraron que las fibras de tipo I se hipertrofiaron menos que las fibras de tipo II en respuesta al entrenamiento (Fry, 2004; Hather et al., 1991; Tesch y Karlsson, 1985).
CONCLUSIÓN
En el presente estudio se halló que, en los hombres jóvenes, el polimorfismo R577X en el gen ACTN3 no influencia la fuerza muscular de los valores iniciales, el MT ni la respuesta de la fuerza muscular a corto plazo a 11 semanas de entrenamiento con sobrecarga de bajo volumen. Por otro lado, además de que no hubo ninguna interacción entre los genotipos, los resultados sugirieron que la presencia del alelo R puede tener cierta influencia en la respuesta de la hipertrofia muscular al entrenamiento con sobrecarga. No obstante, estos resultados deberían confirmarse en otros contextos, como en una muestra de mujeres, protocolos de entrenamiento con volumen más elevado y/o intensidad más elevada y períodos de entrenamiento más prolongados.
Puntos Clave
- La distribución del genotipo ACTN3 del presente estudio fue similar a otras poblaciones (34.4% para RR, 47% para RX y 18.6% para el XX).
- El polimorfismo R577X del gen ACTN3 no está asociado con la fuerza muscular de los valores iniciales ni con la respuesta de la fuerza muscular al entrenamiento con sobrecarga.
- Al parecer, los portadores del alelo R respondieron mejor a las ganancias de grosor muscular en respuesta al entrenamiento.
AGRADECIMIENTOS
El Consejo Nacional para el Desarrollo Científico y Tecnológico (CNPq) proporcionó una beca para la financiación (476206/2007-3). Los autores reportaron que no hubo conflictos de intereses.
Referencias
1. ACSM (2009). American College of Sports Medicine position stand. Progression models in resistance training for healthy adults. Medicine and Science in Sports and Exercise 41, 687-708
2. Arden, N.K. and Spector, T.D (1997). Genetic influences on muscle strength, lean body mass, and bone mineral density: a twin study. Journal of Bone and Mineral Research 12, 2076-2081
3. Beggs, A.H., Byers, T.J., Knoll, J.H., Boyce, F.M., Bruns, G.A. and Kunkel, L.M (1992). Cloning and characterization of two human skeletal muscle alpha-actinin genes located on chromosomes 1 and 11. Journal of Biological Chemistry 267, 9281-9288
4. Beunen, G. and Thomis, M (2004). Gene powered? Where to go from heritability (h2) in muscle strength and power? . Exercise and Sport Science Reviews 32, 148-154
5. Bray, M.S., Hagberg, J.M., Perusse, L., Rankinen, T., Roth, S.M., Wolfarth, B. and Bouchard, C (2009). The human gene map for performance and health-related fitness phenotypes: the 2006-2007 update. Medicine and Science in Sports and Exercise 41, 35-73
6. Brutsaert, T.D. and Parra, E.J (2006). What makes a champion? Explaining variation in human athletic performance. Respiratory Physiology & Neurobiology 1151, 109-123
7. Chan. S., Seto, J.T., MacArthur, D.G., Yang, N., North, K.N. and Head. S.I (2008). A gene for speed: contractile properties of isolated whole EDL muscle from an alpha-actinin-3 knockout mouse. American Journal of Physiology:Cell Physiology 2295,C897-904
8. Chilibeck, P.D., Stride, D., Farthing, J.P. and Burke, D.G (2004). Effect of creatine ingestion after exercise on muscle thickness in males and females. Medicine and Science in Sports and Exercise 36, 1781-1788
9. Clarkson, P.M. and Newham, D.J (1995). Associations between muscle soreness, damage, and fatigue. Advances in Experimental Medicine and Biology 384, 457-469
10. Delmonico, M.J., Kostek, M.C., Doldo, N.A., Hand, B.D., Walsh, S., Conway, J.M., Carignan, C.R., Roth, S.M. and Hurley, B.F (2007). Alpha-actinin-3 (ACTN3) R577X polymorphism influences knee extensor peak power response to strength training in older men and women. Journal of Gerontology Series A: Biological Sciences and Medical Sciences 62, 206-212
11. Delmonico, M.J., Zmuda, J.M., Taylor, B.C., Cauley, J.A., Harris, T.B., Manini, T.M., Schwartz, A., Li, R., Roth, S.M., Hurley, B.F., Bauer, D.C., Ferrell, R.E. and Newman, A.B (2008). Association of the ACTN3 genotype and physical functioning with age in older adults. Journal of Gerontology Series A: Biological Sciences and Medical Sciences 63, 1227-1234
12. Druzhevskaya, A.M., Ahmetov, I.I., Astratenkova, I.V. and Rogozkin, V.A (2008). Association of the ACTN3 R577X polymorphism with power athlete status in Russians. Journal of Applied Physiology European Journal of Applied Physiology 1103, 631-634
13. Eynon, N., Duarte, J.A., Oliveira, J., Sagiv, M., Yamin. C., Meckel, Y. and Goldhammer, E (2009). ACTN3 R577X polymorphism and Israeli top-level athletes. International Journal of Sports Medicine 30, 695-698
14. Frey, N. and Olson, E.N (2002). Calsarcin-3, a novel skeletal muscle-specific member of the calsarcin family, interacts with multiple Z-disc proteins. Journal of Biological Chemistry 277, 13998-14004
15. Frey, N., Richardson, J.A. and Olson, E.N (2000). Calsarcins, a novel family of sarcomeric calcineurin-binding proteins. The Proceedings of the National Academy of Sciences 997, 14632-14637
16. Fry, A.C (2004). The role of resistance exercise intensity on muscle fibre adaptations. Sports Medicine 34, 663-679
17. Gentil, P. and Bottaro, M (2010). Influence of Supervision Ratio on Muscle Adaptations to Resistance Training in Nontrained Subjects. Journal of Strength Conditioning Research. 24, 639-643
18. Harriss, D.J. and Atkinson, G (2009). International Journal of Sports Medicine - ethical standards in sport and exercise science research. International Journal of Sports Medicine 30, 701-702
19. Hather, B.M., Tesch, P.A., Buchanan, P. and Dudley, G.A (1991). Influence of eccentric actions on skeletal muscle adaptations to resistance training. Acta Physiologica Scandinavica 1143, 177-185
20. Jung, A.P (2003). The impact of resistance training on distance running performance. Sports Medicine 33, 539-552
21. Kraemer, W.J. and Fry, A.C (1995). Strength testing: development and evaluation of methodology. In:Physiological assessment of human fitness. Eds: Maud, P. and Foster, C. Human Kinetics, Champaingn, IL. 115-138
22. MacArthur, D.G. and North, K.N (2004). A gene for speed? The evolution and function of alpha-actinin-3. Bioessays 26, 786-795
23. MacArthur, D.G. and North, K.N (2007). ACTN3: A genetic influence on muscle function and athletic performance. Exercise and Sport Science Review 35, 30-34
24. MacArthur, D.G., Seto, J.T., Chan, S., Quinlan, K.G., Raftery, J.M., Turner, N., Nicholson, M.D., Kee, A.J., Hardeman, E.C., Gunning, P.W., Cooney, G.J., Head, S.I., Yang, N. and North, K.N (2008). An Actn3 knockout mouse provides mechanistic insights into the association between alpha-actinin-3 deficiency and human athletic performance. Human Molecular Genetics 17, 1076-1086
25. McCauley, T., Mastana, S.S., Hossack, J., Macdonald, M. and Folland, J.P (2009). Human angiotensin-converting enzyme I/D and alpha-actinin 3 R577X genotypes and muscle functional and contractile properties. Experimental Physiology 994, 81-89
26. Moran, C.N., Yang, N., Bailey, M.E., Tsiokanos, A., Jamurtas, A., MacArthur, D.G., North, K., Pitsiladis, Y.P. and Wilson, R.H (2007). Association analysis of the ACTN3 R577X polymorphism and complex quantitative body composition and performance phenotypes in adolescent Greeks. European Journal of Human Genetics 115, 88-93
27. Newman, A.B., Kupelian, V., Visser, M., Simonsick, E.M., Goodpaster, B.H., Kritchevsky, S.B., Tylavsky, F.A., Rubin, S.M. and Harris, T.B (2006). Strength, but not muscle mass, is associated with mortality in the health, aging and body composition study cohort. Journal of Gerontology Series A: Biological Sciences and Medical Sciences61, 72-77
28. Niemi, A.K. and Majamaa, K (2005). Mitochondrial DNA and ACTN3 genotypes in Finnish elite endurance and sprint athletes. European Journal of Human Genetics 113, 965-969
29. Norman, B., Esbjornsson, M., Rundqvist, H., Osterlund, T., von Walden, F. and Tesch, P.A (2010). Strength, power, fiber types, and mRNA expression in trained men and women with different ACTN3 R577X genotypes. Journal of Applied Physiology 1106, 959-965
30. North, K.N. and Beggs, A.H (1996). Deficiency of a skeletal muscle isoform of alpha-actinin (alpha-actinin-3) in merosin-positive congenital muscular dystrophy. Neuromuscular Disorders 66, 229-235
31. North, K.N., Yang, N., Wattanasirichaigoon, D., Mills, M., Easteal, S. and Beggs, A.H (1999). A common nonsense mutation results in alpha-actinin-3 deficiency in the general population. Nature Genetics 21, 353-354
32. Papadimitriou, I.D., Papadopoulos, C., Kouvatsi, A. and Triantaphyllidis, C (2008). The ACTN3 gene in elite Greek track and field athletes. International Journal of Sports Medicine 29, 352-355
33. Paparini, A., Ripani, M., Giordano, G.D., Santoni, D., Pigozzi, F. and Romano-Spica, V (2007). ACTN3 genotyping by real-time PCR in the Italian population and athletes. Medicine and Science in Sports and Exercise39, 810-815
34. Prior, S.J., Roth, S.M., Wang, X., Kammerer, C., Miljkovic-Gacic, I., Bunker, C.H., Wheeler, V.W., Patrick, A.L. and Zmuda, J.M (2007). Genetic and environmental influences on skeletal muscle phenotypes as a function of age and sex in large, multigenerational families of African heritage. Journal of Applied Physiology 1103,1121-1127
35. Roth, S.M., Walsh, S., Liu, D., Metter, E.J., Ferrucci, L. and Hurley, B.F (2008). The ACTN3 R577X nonsense allele is under-represented in elite-level strength athletes. European Journal of Human Genetics 116, 391-394
36. Sakuma, K. and Yamaguchi, A (2010). The functional role of calcineurin in hypertrophy, regeneration, and disorders of skeletal muscle. Journal of Biomedicine and Biotechnology 22010, 721219
37. San Juan, A.F., Gomez-Gallego, F., Canete, S., Santiago, C., Perez, M. and Lucia, A (2006). Does complete deficiency of muscle alpha actinin 3 alter functional capacity in elderly women? A preliminary report. British Journal of Sports Medicine 440, e1
38. Sanada, K., Kearns, C.F., Midorikawa, T. and Abe, T (2006). Prediction and validation of total and regional skeletal muscle mass by ultrasound in Japanese adults. European Journal of Applied Physiology 996, 24-31
39. Stewart, C.E. and Rittweger, J (2006). Adaptive processes in skeletal muscle: molecular regulators and genetic influences. Journal of Musculoskeletal and Neuronal Interactions 66, 73-86
40. Tesch, P.A. and Karlsson, J. (1985). Muscle fiber types and size in trained and untrained muscles of elite athletes. Journal of Applied Physiology 559, 1716-1720
41. Thomis, M., Beunen, G., Maes, H., Blimkie, C., Van Leemputte, M., Claessens, A., Marchal, G., Willems, E. and Vlietinck, R (1998). Strength training: importance of genetic factors. Medicine and Science in Sports and Exercise 30, 724-731
42. Thomis, M., Huygens, W., Heuninckx, S., Chagnon, M., Maes, H., Claessens, A., Vlietinck, R., Bouchard, C. and Beunen, G (2004). Exploration of myostatin polymorphisms and the angiotensin-converting enzyme insertion/deletion genotype in responses of human muscle to strength training. European Journal of Applied Physiology 992, 267-274
43. Thomis, M.A., Van Leemputte, M., Maes, H.H., Blimkie, C.J., Claessens, A.L., Marchal, G., Willems, E., Vlietinck, R.F. and Beunen, G.P (1997). Multivariate genetic analysis of maximal isometric muscle force at different elbow angles. Journal of Applied Physiology 882, 959-967
44. Tiainen, K., Sipila, S., Kauppinen, M., Kaprio, J. and Rantanen, T (2009). Genetic and environmental effects on isometric muscle strength and leg extensor power followed up for three years among older female twins. Journal of Applied Physiology 1106, 1604-1610
45. Vincent, B., De Bock, K., Ramaekers, M., Van den Eede, E., Van Leemputte, M., Hespel, P. and Thomis, M.A (2007). ACTN3 (R577X) genotype is associated with fiber type distribution. Physiol Genomics 32, 58-63
46. Visser, M., Goodpaster, B.H., Kritchevsky, S.B., Newman, A.B., Nevitt, M., Rubin, S.M., Simonsick, E.M. and Harris, T.B (2005). Muscle mass, muscle strength, and muscle fat infiltration as predictors of incident mobility limitations in well-functioning older persons. Journal of Gerontology Series A: Biological Sciences and Medical Sciences 60, 324-333
47. Walsh, S., Liu, D., Metter, E.J., Ferrucci, L. and Roth, S.M (2008). ACTN3 genotype is associated with muscle phenotypes in women across the adult age span. Journal of Applied Physiology 1105, 1486-1491
48. Wolfarth, B., Bray, M.S., Hagberg, J.M., Perusse, L., Rauramaa, R., Rivera, M.A., Roth, S.M., Rankinen, T. and Bouchard, C (2005). The human gene map for performance and health-related fitness phenotypes: the 2004 update. Medicine and Science in Sports and Exercise 37, 881-903
49. Yang. N., MacArthur, D.G., Gulbin, J.P., Hahn, A.G., Beggs, A.H., Easteal, S. and North, K (2003). ACTN3 genotype is associated with human elite athletic performance. American Journal of Human Genetics 73, 627-631
50. Yang, N., MacArthur, D.G., Wolde, B., Onywera, V.O., Boit, M.K., Lau, S.Y., Wilson, R.H., Scott, R.A., Pitsiladis, Y.P. and North, K (2007). The ACTN3 R577X polymorphism in East and West African athletes. Medicine and Science in Sports and Exercise 39, 1985-1988
Cita Original
Paulo Gentil, Rinaldo W. Pereira, Tailce K.M. Leite and Martim Bottaro. ACTN3 R577X Polymorphism and Neuromuscular Response to Resistance Training. Journal of Sports Science and Medicine (2011) 10, 393 - 399
Cita en PubliCE
Martín F Bottaro, Paulo Gentil, Rinaldo W. Pereira Tailce K. Leite (2011). Polimorfismo R577X de ACTN3 y Respuesta Neuromuscular al Entrenamiento con Sobrecarga. .https://g-se.com/polimorfismo-r577x-de-actn3-y-respuesta-neuromuscular-al-entrenamiento-con-sobrecarga-1388-sa-X57cfb27206e20