Mejora de la Eficiencia Muscular Desarrollada a Medida que Madura un Campeón del Tour de Francia
Edward F Coyle11 Human Performance Laboratory, Department of Kinesiology and Health Education, The University of Texas at Austin, TX 78712, Estados Unidos.
Resumen
Este estudio de caso describe la maduración fisiológica desde los 21 a los 28 años del ciclista que se ha convertido recientemente en el seis veces consecutivas Gran Campeón del Tour de Francia, a las edades de 27-32 años. El máximo consumo de oxígeno (VO2 máx.) en el estado entrenado permaneció en ~6 L/min, la masa corporal magra permaneció en ~70 kg, y la frecuencia cardíaca máxima declino desde 207 hasta 200 lat./min. El umbral del lactato fue el típico de un ciclista de nivel competitivo y se produjo al 76-85 % del VO2 máx., sin embargo la concentración de lactato sanguíneo máxima fue notablemente baja en el estado entrenado. Parece que una mejora del 8% en la eficiencia muscular y así en la producción de potencia a un dado consumo de oxígeno (VO2) es la característica que más mejoró a medida que el atleta maduró desde la edad de 21 a 28 años. Es importante destacar que a la edad de 25 años, este campeón desarrolló un cáncer avanzado, que requirió cirugías y quimioterapia. Durante los meses que condujeron a cada una de sus victorias en el Tour de Francia, el ciclista redujo su masa corporal y su grasa corporal en 4-7 kg (i.e., ~ 7%). De este modo, a través del período de 7 años, un incremento en la eficiencia muscular y una reducción en la grasa corporal contribuyeron igual a una notable mejora del 18% en su potencia en estado estable por kilogramo de masa corporal cuando pedaleaba a un dado VO2 (e.g., 5 l/min). Es posible plantear la hipótesis que señala que la mejora en la eficiencia muscular probablemente refleja cambios en el tipo de miosina muscular estimulada a partir de años de entrenamiento intenso durante 3-6 h en la mayoría de los días.
Palabras Clave: máximo consumo de oxígeno, concentración de lactato sanguíneo
INTRODUCCION
Se ha aprendido mucho acerca de los factores fisiológicos que contribuyen a la capacidad de rendimiento de resistencia simplemente describiendo las características de los atletas de resistencia en deportes tales como la carrera de larga distancia, competiciones de ciclismo y esquí de fondo. Los numerosos determinantes fisiológicos de la resistencia han sido organizados en un modelo que integra factores, tales como el máximo consumo de oxígeno (VO2 máx.), el umbral del lactato sanguíneo, y la eficiencia muscular, ya que ha sido hallado que estas son las variables más importantes (7, 8, 15, 21). Un enfoque común ha sido medir estos factores fisiológicos en un dado atleta en un punto en el tiempo durante su carrera deportiva y comparar el perfil de este individuo con el de una población de pares (4, 6, 15, 16, 21). Aunque este enfoque describe las variaciones que existen dentro de una población, el mismo no provee información acerca del grado al cual un dado atleta puede mejorar sus factores fisiológicos específicos determinantes del rendimiento de resistencia con años de entrenamiento continuo a medida que el atleta madura y alcanza su potencial fisiológico. Hay notablemente pocos reportes longitudinales que documenten los cambios en factores fisiológicos que acompañan a años de entrenamiento de resistencia continuo al nivel realizado por atletas de resistencia de élite.
Este estudio de caso reporta los cambios fisiológicos que ocurren en un ciclista de nivel competitivo durante un período de desarrollo de 7 años, desde los 21 a los 28 años. La descripción de esta persona es digna de mención por dos razones. En primer lugar, el ciclista llegó a convertirse en el seis veces y actual Gran Campeón del Tour de Francia, y así fueron identificadas las adaptaciones relevantes para esta prueba. Notablemente, el ciclista logró esto luego de desarrollar y recibir tratamiento para un cáncer avanzado. De este modo, este reporte es también importante porque da información, aunque limitada, respecto de la recuperación de la “fisiología del rendimiento” luego de un tratamiento exitosos para un cáncer avanzado. El enfoque de este estudio va a ser reportar los resultados de evaluaciones estandarizadas de laboratorio de este individuo en cinco puntos de tiempo correspondientes a las edades de 21,1, 21,5, 22,0, 25,9 y 28,2 años.
METODOS
Secuencia General de Evaluación
Una vez que el ciclista se reportaba en el laboratorio, eran registrados el historial de entrenamiento, competición y médico, se medía la masa corporal (±0,1 kg), y los test siguientes fueron realizados luego de que fuera obtenido un informe de consentimiento, con procedimientos aprobados por el Comité de Revisión Interna de la Universidad de Texas en Austin. La eficiencia mecánica y el umbral del lactato (LT) fueron determinados en una bicicleta ergométrica estacionaria en un test de 25 min, con incrementos en la tasa de trabajo progresivos cada 5 min en un intervalo de 50, 60, 70, 80 y 90% del VO2 máx. Luego de un período de recuperación activa de 10-20 min, era medido el VO2 máx. del ciclista. Luego era determinada la composición corporal a través de pesaje hidrostático y/o medición del grosor de los pliegues cutáneos (34, 35).
Medición del VO2 máx
El mismo ergómetro Monark (modelo 819) equipado con un asiento de competición y manubrio de carretera y pedales para zapatillas de ciclismo fue usado para todas las evaluaciones de ciclismo, y la altura del asiento y la posición del mismo fueron mantenidas constantes. La longitud de las palancas o bielas era de 170 mm. El VO2 máx. fue medido durante ciclismo continuo que duró entre 8 y 12 min, con incrementos de la tasa de trabajo cada 2 min. Siempre se producía una estabilización del consumo de oxígeno (VO2), y este individuo pedaleaba hasta el agotamiento a una producción de potencia final que fue un 10-20% mayor a la producción de potencia mínima para producir el VO2 máx. Una muestra de sangre venosa era obtenida cada 3-4 min luego del agotamiento para determinar la concentración de lactato sanguíneo después del ejercicio máximo, tal como se describe abajo. El sujeto respiraba a través de una válvula de Daniels; los gases espirados fueron muestreados de manera continua desde una cámara de mezclado y analizados para O2 (Applied Electrochemistry S3A) y CO2 (Bechman LB-2). Los volúmenes de aire inspirado fueron medidos usando un medidor de gas seco (Parkinson-Cowan CD4). Estos instrumentos estaban conectados a través de interfases con una computadora que calculó el VO2 cada 30 s. El mismo equipo fue usado para la calorimetría indirecta a través del período de 7 años, siendo los analizadores de gases calibrados con los mismos gases de concentración conocida y el medidor de gas seco fue calibrado periódicamente con un espirómetro Tissot de 350 litros.
LT Sanguíneo
Los sujetos pedalearon en la bicicleta ergométrica Monark (modelo 819) en forma continua durante 25 min a tasas de trabajo que produjeron ~ 50, 60, 70, 80 y 90 del VO2 máx. para cada etapa sucesiva de 5 min. El ergómetro calibrado fue programado en el modo de potencia constante, y el sujeto mantuvo una cadencia de pedaleo de 85 rpm. Las muestras sanguíneas fueron obtenidas ya sea de un catéter en la vena antecubital o de la punta de los dedos durante el 5to minuto de ejercicio en cada etapa o 4 min después del ejercicio. La sangre fue desproteinizada en ácido perclórico y luego analizada para el lactato usando un método enzimático y espectrofotométrico (20). El LT sanguíneo fue determinado, tal como fue previamente descrito (12), graficando la relación de la concentración de lactato vs. el VO2 y determinando el VO2 en el cual el lactato sanguíneo se incrementó por encima de 1 mM con respecto a la condición inicial. La máxima concentración de lactato sanguíneo fue determinada a partir de una muestra de sangre obtenida durante el 4to minuto después de la determinación del VO2 máx.
Eficiencia Mecánica
La eficiencia general o gross efficiency fue calculada como el cociente entre el trabajo realizado por minuto (i.e., vatios convertidos a kcal/min) y la energía gastada por minuto (kcal/min). El gasto energético por minuto (i.e., kcal/min) fue calculado a partir del VO2 y el índice de intercambio respiratorio, usando tablas de Lusk (31). En un dado día de evaluación, la eficiencia general fue usualmente similar a todas las tasas de trabajo evaluadas cuando el ciclista trabajaba al 50-90% del VO2 máx. y a 80-90 rpm, tal como ha sido previamente descrito en ciclistas entrenados (10, 31). De este modo, la eficiencia general fue reportada como los valores promedio obtenidos a las cinco tasas de trabajo (10).
El delta de eficiencia es definido como el cociente entre el cambio en la tasa de trabajo realizada por minuto y el cambio en el gasto energético por minuto (10, 31). El delta de eficiencia fue identificado a partir de una regresión lineal (y=mx+b) de la relación (i.e., 5 puntos a ~ 50, 60, 70, 80 y 90% del VO2 máx.) entre la energía gastada por minuto (i.e., y; kcal/min) vs. el trabajo realizado por minuto (i.e., x; kcal/min). El delta de eficiencia fue calculado a partir de la pendiente de la relación y fue igual a la recíproca de m (i.e., 1/m) (31).
Composición Corporal
La densidad corporal fue determinada a partir de pesaje hidrostático, con medición directa del volumen residual en los pulmones usando la técnica de dilución de nitrógeno (34, 35). Además, fueron determinados los pliegues cutáneos en 5 sitios, y la suma de estas mediciones fue relacionada con la densidad corporal. Los porcentajes de grasa corporal y el peso corporal magro fueron calculados a partir de la densidad corporal y el peso corporal (35).
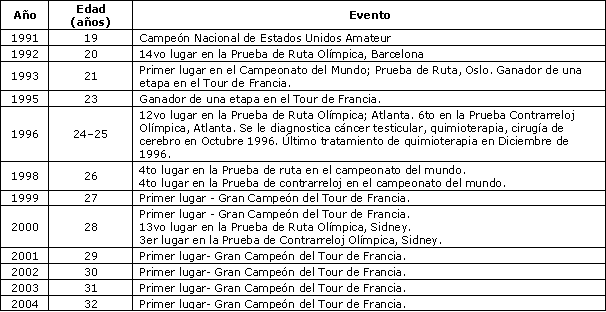
Tabla 1. Resumen de la historia de competiciones ciclísticas e historia médica
del sujeto.
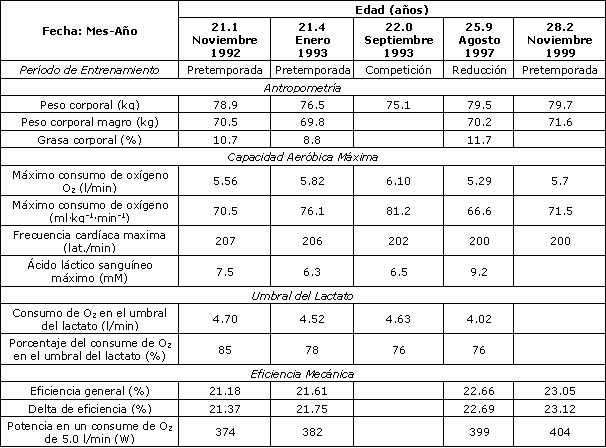
Tabla 2. Características fisiológicas del ciclista desde la edad de 21 a 28
años.
RESULTADOS
Antropometría
El peso corporal total durante una evaluación de laboratorio estuvo entre ~ 76 y 80 kg desde 1992 hasta 1997, así como durante la pretemporada de 1999. Sin embargo, cuando el ciclista compitió en el Tour de Francia en 1999-2004, el peso corporal reportado por el sujeto fue de ~ 72-74 kg. El peso corporal magro fue de ~ 70 kg durante el período de 1992-1997 (Tabla 2). La talla fue de ~ 178 cm.
VO2 máx., Frecuencia Cardíaca Máxima y LT Sanguíneo.El VO2 máx. durante los meses de pretemporada de Noviembre a Enero estuvo entre 5,56 y 5,82 l/min durante el período de 1992-1999. El VO2 máx. durante la temporada de 1993, justo después de ganar el Campeonato Mundial de Ciclismo de Ruta (Septiembre de 1993) fue de 6,1 l/min y 81,2 ml.kg-1.min-1, resultados que fueron corroborados por el Comité Olímpico de Estados Unidos (Colorado Springs, CO). Ocho meses después de un tratamiento de quimioterapia para el cáncer y durante un período de entrenamiento inconsistente y reducido (i.e., Agosto de 1997), el VO2 máx. fue de 5,29 l/min y 66,6 ml.kg-1.min-1. Además, en este tiempo de entrenamiento reducido, la máxima concentración de lactato medida 4 min después del agotamiento fue 9,2 mM en comparación con los valores previamente registrados entre 6,3-7,5 mM. La frecuencia cardíaca máxima disminuyó desde 207 a 200 lat./min desde 1992 hasta 1999. El VO2 correspondiente al umbral del lactato fue 4,5-4,7 l/min, cuando fue medido en 1992-1993 y, tal como se esperaba, se redujo a 4,02 l/min durante el período de entrenamiento reducido en Agosto de 1997.
Eficiencia Mecánica
Los valores de eficiencia general y el delta de eficiencia durante el período de 1992 hasta 1999 son presentados en la Figura 1. Este incremento progresivo en la eficiencia constituyó un incremento de 8-9% a través de este período. Esta mejora es también evidenciada en la medición de la potencia generada cuando el ciclista pedalea a un VO2 dado de 5,0 l/min, y este valor se incrementó desde 374 hasta 403 W (i.e., 8%; Tabla 2). Dado que el éxito en el Tour de Francia es típicamente determinado cuando se pedalea cuesta arriba en las montañas, es mejor normalizar la potencia con el peso corporal (i.e., W/kg). Dada la reducción del peso corporal del individuo desde 78,9 kg (en 1992) hasta ~ 72 kg durante sus victorias en el Tour de Francia y dado su incremento en la eficiencia muscular, se calcula que su índice potencia/masa corporal (i.e., potencia/kg) cuando pedaleaba a 5,0 l/min se incrementó un notable 18% desde 1992 hasta 1999 (i.e., 4,74 vs. 5,60 W/kg cuando el VO2 estaba en 5,0 l/min). Teniendo en cuenta que su VO2 máx. se mantuvo en ~ 6 l/min, este VO2 de 5,0 l/min representa ~ 83% del VO2 máx. De este modo, su “potencia por kilogramo” a un dado porcentaje del VO2 máx. (e.g., 83%) se incrementó en un 18%.
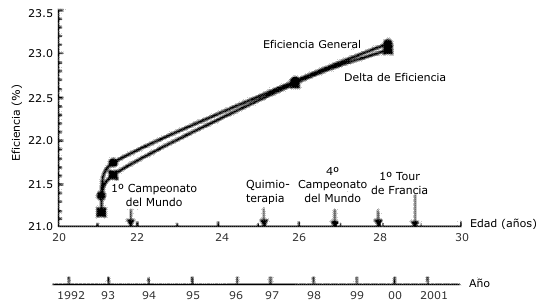
Figura 1. Eficiencia mecánica cuando el ciclista pedalea expresada como
“eficiencia general” y “delta de eficiencia” a través de un período de 7 años en
la carrera deportiva de este individuo. Tour de Francia: Gran Campeón del Tour
desde 1999-2004.
DISCUSION
Este estudio de caso ha descrito las características fisiológicas de un renombrado ciclista de ruta competitivo, quien es actualmente el seis veces Gran Campeón del Tour de Francia. El estudio reporta que el factor fisiológico más relevante para la mejora del rendimiento a medida que el ciclista maduró a través de un período de 7 años, desde las edades de 21 a 28 años, fue un incremento en la eficiencia muscular del 8% mientras el ciclista pedalea. Esta adaptación combinada con reducciones relativamente grandes en la grasa corporal y así en el peso corporal (e.g., 78-72 kg) durante los meses antes del Tour de Francia contribuyeron a un incremento impresionante del 18% en el índice potencia/peso corporal del ciclista (i.e., W/kg) cuando el ciclista pedaleaba a un dado VO2 (e.g., 5 l/min o ~ 83% del VO2 máx.). Notablemente, este individuo fue capaz de lograr estas mejoras a pesar del hecho de haber desarrollado un cáncer avanzado a la edad de 25 años y haber requerido cirugías y quimioterapia.
En el estado entrenado, el individuo posee un notablemente alto VO2 máx. de ~ 6 l/min y su LT sanguíneo ocurrió a un VO2 de 4,6 l/min (i.e., 76-85% del VO2 máx.). Estos factores fisiológicos permanecieron relativamente estables desde las edades de 21 a 28 años. Estos valores absolutos son más altos que los que hemos medido en ciclistas de competición a nivel nacional en Estados Unidos (9), varios de los cuales han competido subsiguientemente a nivel profesional en Europa durante el período de 1989-1995. Ha sido reportado que el 5 veces gran Campeón del Tour de Francia durante los años 1991-1995 posee un VO2 máx. de 6,4 l/min y 79 ml.kg-1.min-1 con un peso corporal de 81 kg (28). Las mediciones de laboratorio del sujeto de nuestro estudio no fueron realizadas justo después del Tour de Francia; sin embargo, con la idea prudente acerca de que el VO2 máx. fue de al menos 6,1 l/min y dado que este sujeto reportó un peso corporal de 72 kg, nosotros estimamos que su VO2 máx. debe haber sido de 85 ml.kg-1.min-1 durante el período de sus victorias en el Tour de Francia. De este modo, su VO2 máx. por kilogramo de peso corporal durante sus victorias de 1999-2004 parece ser de algún modo mayor al que fue reportado para el campeón durante 1991-1995 y parece estar entre los valores más altos reportados para corredores y ciclistas de clase mundial (e.g., 80-85 ml.kg-1.min-1) (6, 15, 16, 28, 29).
Es generalmente apreciado que además de un alto VO2 máx., el éxito en deportes de resistencia también requiere una capacidad para ejercitarse por períodos de tiempo prolongados a altos porcentajes del VO2 máx., así como la capacidad de convertir esa energía (i.e., ATP) en potencia muscular y velocidad (5, 7, 8, 29). La identificación del LT sanguíneo (e.g., incremento de 1 mM en el lactato sanguíneo por encima de la línea de base) en términos absolutos o como porcentaje del VO2 máx., es, en si mismo, una variable de predicción razonable del rendimiento aeróbico (i.e., tiempo que puede ser mantenida una determinada tasa de recambio o turnover de ATP) (7, 8, 14, 21), y la predicción es fortalecida aún más cuando se combina la medición de la densidad capilar muscular con el LT (11). Se piensa que la densidad capilar es un índice de la capacidad de los músculos que trabajan de limpiar los metabolitos fatigantes (e.g., ácido) de las fibras musculares hacia la circulación, mientras que se piensa que el LT refleja la producción de metabolitos fatigantes en las fibras musculares (7, 8).
Como se esperaba, este individuo posee un alto LT en un intervalo de 76-85% del VO2 máx. Sin embargo, el aspecto único del perfil del lactato sanguíneo de este individuo fue su extremadamente baja concentración de lactato medida 4 min después del agotamiento durante la medición del VO2 máx. En este sujeto el lactato sanguíneo máximo en el estado entrenado fue de solo 6,5-7,5 mM. Por comparación, todos los ciclistas competitivos que hemos evaluados, incluyendo los compañeros de equipo de este sujeto, poseen niveles de lactato sanguíneo máximo post-ejercicio en un intervalo de 9-14 mM (9, 11). El mecanismo de esta máxima concentración de lactato sanguíneo extremadamente baja no está claro, aunque probablemente refleje una reducida producción de lactato cuando el sujeto se ejercita hasta el agotamiento a intensidades por encima del VO2 máx. Una posibilidad es que la actividad de las enzimas musculares responsables en gran medida de la producción de lactato [i.e., lactato deshidrogenasa (LDH) y fosforilasa] estén fuertemente atenuadas en este individuo cuando está entrenado (12, 24). Debería ser señalado que este individuo en vez de agotarse durante la evaluación del VO2 máx., presenta el patrón característico del ciclista competitivo, incluyendo una meseta o plateau en el VO2 y la frecuencia cardíaca en valores máximos por 1-3 min, hiperventilación moderada, índice de intercambio respiratorio >1,05 y una pérdida progresiva de la cadencia de pedaleo a una potencia constante durante 30-60 s antes del agotamiento.
Es interesante señalar que cuando el VO2 máx. y la concentración de lactato sanguíneo fueron medidos durante el período de entrenamiento reducido 8 meses después de la quimioterapia (25,9 años de edad), la máxima concentración de lactato sanguíneo se incrementó hasta 9,2 mM. Esto coincide con nuestras observaciones previas que indican que el desentrenamiento incrementa la máxima concentración de lactato sanguíneo en atletas de resistencia bien entrenados desde 10 hasta 12,5 mM en asociación con un incremento de 21% en la actividad total de la LDH (i.e., 21%) (12, 13, 24). Debería ser señalado que la concentración de lactato sanguíneo fue medida, en este estudio y en otros estudios previos, exclusivamente con un análisis espectrofotométrico del NADH producido a partir de la reacción de la LDH después de completar el lisado de las células rojas sanguíneas (20). Teniendo en cuenta nuestra experiencia, las concentraciones máximas de lactato medidas usando analizadores automáticos disponibles comercialmente son menores, posiblemente debido al lisado incompleto de las células rojas sanguíneas a pesar de la adición del detergente a los reactantes (3).
La evaluación fisiológica fue realizada 8 meses después de la quimioterapia durante un período de entrenamiento reducido. Con respecto a su entrenamiento previo, durante el 3er y 4to mes después de la quimioterapia, el ciclista entrenó ~ 5 días/semana durante 2-5 horas a una intensidad moderada. Durante el 5to y 6to mes, la intensidad del entrenamiento disminuyó. Durante el período del 6to y 7to mes después de la quimioterapia, el ciclista no realizó entrenamiento de resistencia. Sin embargo, durante los 8 días anteriores a la evaluación fisiológica de laboratorio (i.e., 8 meses después de la quimioterapia), el ciclista entrenó 1-2 h/días a una intensidad moderada, a frecuencias cardíacas entre 120-150 latidos/min. Durante esta evaluación de laboratorio 8 meses después de completar la quimioterapia, este individuo no presentó efectos de la enfermedad de sus cirugías y quimioterapia previas. En particular, el volumen ventilatorio durante el ejercicio máximo pareció ser el típico, y sus respuestas cardiovasculares fueron normales a frecuencias cardíacas de 120-150 lat./min. Además, la frecuencia cardíaca máxima alcanzó el nivel saludable para este individuo (i.e., 200 lat./min). Sin embargo, tal como se esperaba debido a la reducción del entrenamiento, el VO2 máx. fue un 6-12% menor con un valor de 5,3 l/min y 67 ml.kg-1.min-1. Si este individuo no realizara entrenamiento durante 3 meses, nosotros predecimos que su VO2 máx. se estabilizaría en 5 l/min (e.g., 61-63 ml.kg-1.min-1 para una masa corporal de 80 kg) en base a nuestras mediciones previas en atletas de resistencia bien entrenados durante el desentrenamiento (13; ver Figura 1). En general se cree que un VO2 máx. de 56-62 ml.kg-1.min-1 es el valor más alto que puede alcanzar un hombre promedio que no está dotado genéticamente para la resistencia con entrenamiento de resistencia prolongado y muy intenso (13, 23). Por lo tanto, parece que en el estado desentrenado, el VO2 máx. de este individuo está en el intervalo de los valores más altos que un hombre normal puede alcanzar con el entrenamiento.
Los mecanismos fisiológicos responsables del incremento del 8%, en tanto la eficiencia general como el delta de eficiencia durante el ciclismo, así como el estímulo que provoca esta adaptación, no están claros. La observación de que tanto la eficiencia general como el delta de eficiencia mejoran en un mismo grado y también en el mismo transcurso de tiempo (Figura 1) sugiere una mejora de la eficiencia del recambio o turnover de ATP dentro de las fibras musculares durante la contracción (10, 31). Esto se debe a que la medición del delta de eficiencia, definida como el incremento en la producción de potencia relativa al incremento en el gasto energético (calculado a partir del VO2) a través de un amplio intervalo de tasas de trabajo, proporciona el mejor reflejo de la producción de potencia a partir de la formación de los puentes cruzados de actina y miosina en los músculos activos (26), ya que elimina o minimiza la influencia del costo energético del ciclismo sin carga, el trabajo ventilatorio y otros procesos metabólicos no directamente relacionados a la producción de potencia muscular (31). Nosotros reportamos previamente, a partir de observaciones transversales de ciclistas competitivos, que el porcentaje de fibras musculares tipo I del vasto lateral está directa y positivamente relacionado tanto al delta de eficiencia como a la eficiencia general medida tanto durante el ciclismo como durante una tarea simple de extensión de rodilla (10, 25). De este modo, un posible mecanismo para el incremento en la eficiencia es que este individuo incrementó su porcentaje de fibras musculares tipo I durante este período de estudio de 7 años.
Usando nuestra predicción previamente reportada acerca del porcentaje de fibras musculares tipo I a partir de mediciones directas de eficiencia general y mecánica en este individuo, predecimos que el podría haber incrementado su porcentaje de fibras musculares tipo I desde 60 a 80%. De manera interesante, esta magnitud de incremento en las fibras tipo I con 7 años de entrenamiento de resistencia continuo en este individuo es notablemente similar a nuestra predicción hecha en 1991 en base a observaciones transversales de ciclistas competitivos (9; ver Figura 8). Para nuestro conocimiento, no ha habido estudios longitudinales realizados a través de los años en humanos, que hayan evaluado directamente la hipótesis que plantea que las fibras tipo II pueden ser convertidas a fibras musculares tipo I con el entrenamiento de resistencia continuo e intenso. Sin embargo, durante períodos de entrenamiento de resistencia extremo en ratas, parece que el músculo esquelético sufre conversión de fibras tipo II a tipo I (18). Otros factores que se ha reportado, incrementan la eficiencia en el ciclismo y la economía de carrera, son la exposición intermitente a la hipoxia durante varias semanas, como la que realizan los atletas que pasan períodos viviendo a alturas elevadas o en ambiente hipóxicos (19, 30). Tal como muchos atletas de resistencia, este individuo ha incorporado exposiciones hipóxicas en su plan anual, el cual puede ser otro factor que contribuye al incremento de la eficiencia durante el pedaleo.
Ha sido reconocido por décadas que el entrenamiento de resistencia de las ratas, incrementa la actividad miosín-ATPasa de las fibras tipo I mientras que la disminuye en las fibras tipo II (2). Estudios más recientes realizados en humanos por Fitts, Costill y colegas (17, 32, 33) midieron en forma directa la velocidad máxima de acortamiento de fibras musculares aisladas (i.e., usando el slack test) obtenidas a partir de biopsia. Diez semanas de natación de alta intensidad (e.g., 4-5 km/día) incrementaron la velocidad máxima de las fibras tipo I, mientras que en la fibras tipo II, la misma disminuyó (17). Además, Widrick et al. (32, 33) hallaron que sujetos de sexo masculino que realizaron altos niveles de actividad física por 20-25 años y que eran corredores master de elite también presentaron un incremento de la velocidad máxima de las fibras tipo I, el cual fue asociado con un tipo alterado de miosina (i.e., 28% más de miosina de cadena liviana 3 vs. 2). De este modo, el entrenamiento de resistencia intenso realizado por períodos prolongados de tiempo resulta en alteraciones en la actividad miosín-ATPasa, donde las fibras tipo II se vuelven más parecidas a las fibras tipo I y estas últimas incrementan la actividad ATPasa y alteran el tipo de miosina e incrementan la máxima velocidad de acortamiento. Estas observaciones apoyan la posibilidad que en el sujeto del presente estudio, los 7 años de entrenamiento de resistencia extremadamente intenso y el incremento de la eficiencia muscular mientras pedalea, estuvieran relacionados al tipo alterado de miosina que permitió que una mayor cantidad de la energía liberada de la hidrólisis del ATP durante la contracción sea convertida en producción de potencia.
En este atleta no se tomaron muestras musculares obtenidas quirúrgicamente para probar directamente esta hipótesis acerca de que la conversión del tipo de fibra muscular contribuyó al gran incremento en la eficiencia mecánica o muscular mientras se pedalea. De este modo, esta hipótesis que plantea que en este individuo las fibras musculares tipo I se incrementaron, requiere la identificación de otras características de rendimiento que hayan cambiado en este individuo durante el período de 7 años, teniendo en cuenta la discusión de si las mismas son consistentes con la hipótesis del incremento del porcentaje de fibras musculares tipo I.
Aunque durante todas las mediciones de laboratorio de la eficiencia mecánica, la cadencia de pedaleo fue mantenida constante a 85 rpm, la cadencia de este individuo elegida libremente durante una competición en modalidad contrarreloj de 30 a 60 min de duración, se incrementó progresivamente durante este período de 7 años desde ~ 85-95 rpm hasta ~ 105-110 rpm. Efectivamente, este incremento en las revoluciones por minuto elegidas libremente cuando se pedalea a alta intensidad es consistente con los incrementos en las fibras musculares tipo I, debido a que los ciclistas con un alto porcentaje de fibras tipo I eligen una mayor cadencia de pedaleo cuando se ejercitan a altas producciones de potencia (22). Aunque esto inicialmente parece paradójico, mayores cadencias de pedaleo sirven tanto para acercar la velocidad de contracción de las fibras musculares a la velocidad de máxima potencia como para reducir la fuerza muscular y de pedaleo requerida para cada ciclo de pedaleo. Se debe tener en cuenta que cuando se realiza ejercicio a una determinada tasa del metabolismo oxidativo, un incremento del 8% en la eficiencia mecánica va a resultar en un 8% más de desarrollo de potencia y fuerza muscular en los pedales, cuando la cadencia de pedaleo es mantenida constante. A medida que se incrementa la eficiencia de pedaleo debido al incremento del porcentaje de fibras musculares tipo I, es posible que el incremento de la potencia se manifieste incrementando la cadencia de pedaleo (i.e., velocidad) en vez de incrementando la fuerza muscular dirigida a los pedales. Este enfoque parece producir menos sensación de esfuerzo relativo a la fuerza muscular (27). De este modo, es probable que los incrementos en la cadencia de pedaleo elegida libremente desplegados a través de los años por este campeón del Tour de Francia reflejen su incremento en la eficiencia mecánica, coincidiendo con el patrón esperado como resultado de la conversión de fibras musculares desde el tipo II al tipo I.
Este reporte ha identificado el factor fisiológico que mejoró más desde las edades de 21 a 28 años en el ciclista que actualmente se ha convertido en el seis veces consecutivas Gran Campeón del Tour de Francia es la eficiencia muscular. Como resultado de esto, la producción de potencia mientras se pedalea a un VO2 absoluto de 5,0 l/min se incrementó en un 8%. Otro factor que le permitió a este individuo convertirse en Gran Campeón del Tour de Francia fue sus grandes reducciones en el peso corporal y grasa corporal durante los meses previos a la carrera. De este modo, a través del período de 7 años, el ciclista desplegó un incremento notable de 18% en su potencia en estado estable por kilogramo de peso corporal mientras pedaleaba a un dado VO2 (e.g., 5 l/min). Nosotros planteamos la hipótesis que indica que el incremento de la eficiencia muscular podría reflejar alteraciones en el tipo de miosina muscular estimulado a partir de los años de entrenamiento intenso de 3-6 horas durante la mayoría de los días. Es notable que a la edad de 25 años este individuo desarrolló un cáncer avanzado, que requirió cirugías y quimioterapia, sin embargo estos eventos no parecieron impedir que se produjera su maduración fisiológica y logros atléticos. Claramente, este campeón materializa un fenómeno tanto de selección natural genética como el extremo al cual el humano puede adaptarse al entrenamiento de resistencia realizado por una década o más en una persona que está verdaderamente inspirada.
Agradecimientos
El autor aprecia mucho la respetuosa colaboración y actitud positiva de Lance Armstrong a través de los años.
Dirección para el Envío de Correspondencia
E.F. Coyle, Bellmont may 222, Dep.of Kinesiology and Health Education, The Univ. of Texas at Austin, Austin, TX 78712, correo electrónico: coyle@mail.utexas.edu.
Referencias
1. Armstrong L (2000). It is Not About the Bike. New York: Putman
2. Baldwin K, Winder W, and Holloszy JO (1975). Adaptation of actomyosin. ATPase in different types of muscle to endurance exercise. Am J Physiol 229: 422426
3. Bishop P, Smith J, Kime J, Mayo J, and Tin Y (1992). Comparison of a manual and an automated enzymatic technique for determining blood lactate concentrations.. Int J Sports Med 13: 3639
4. Burke E, Cerny F, Costill D, and Fink W (1977). Characteristics of skeletal muscle in competitive cyclists. Med Sci Sports 9: 109112
5. Cavanagh PR, Pollock ML, and Landa J (1977). A biomechanical comparison of elite and good distance runners. Ann NY Acad Sci 301: 328345
6. Costill D, Fink W, and Pollock M (1976). Muscle fiber composition and enzyme activities of elite distance runners. Med Sci Sports 8: 96100
7. Coyle E (1995). Integration of the physiological factors determining endurance performance ability. Exerc Sport Sci Rev 23: 2563
8. Coyle E (1999). Physiological determinants of endurance exercise performance. J Sci Med Sport 2: 181189
9. Coyle E, Feltner M, Kautz S, Hamilton M, Montain S, Baylor A, Abraham L, and Petrek G (1991). Physiological and biomechanical factors associated with elite endurance cycling performance. Med Sci Sports Exerc 23: 93107
10. Coyle E, Sidossis L, Horowitz J, and Beltz J (1992). Cycling efficiency is related to the percentage of type I muscle fibers. Med Sci Sports Exerc 24: 782788
11. Coyle EF, Coggan AR, Hopper MK, and Walters TJ (1988). Determinants of endurance in well-trained cyclists. J Appl Physiol 64: 26222630
12. Coyle EF, Martin WH III, Bloomfield SA, Lowry OH, and Holloszy JO (1985). Effects of detraining on responses to submaximal exercise. J Appl Physiol 59: 853859
13. Coyle EF, Martin WH III, Sinacore DR, Joyner MJ, Hagberg JM, and Holloszy JO (1984). Time course of loss of adaptations after stopping prolonged intense endurance training. J Appl Physiol 57: 18571864
14. Coyle EF, Martin WH, Ehsani AA, Hagberg JM, Bloomfield SA, Sinacore DR, and Holloszy JO (1983). Blood lactate threshold in some well-trained ischemic heart disease patients. J Appl Physiol 54: 1823
15. Farrell P, Wilmore J, Coyle E, Billing J, and Costill D (1979). Plasma lactate accumulation and distance running performance. Med Sci Sports 11: 338344
16. Fink WJ, Costill DL, and Pollock ML (1977). Submaximal and maximal working capacity of elite distance runners. Part II. Muscle fiber composition and enzyme activities. Ann NY Acad Sci 301: 323327
17. Fitts RH, Costill DL, and Gardetto PR (1989). Effect of swim exercise training on human muscle fiber function. J Appl Physiol 66: 465475
18. Green H, Klug G, Reichmann H, Seedorf U, Wiehrer W, and Pette D (1984). Exercise-induced fibre type transitions with regard to myosin, parvalbumin, and sarcoplasmic reticulum in muscles of the rat. Pflügers Arch 400: 432438
19. Green HJ, Roy B, Grant S, Hughson R, Burnett M, Otto C, Pipe A, McKenzie D, and Johnson M (2000). Increases in submaximal cycling efficiency mediated by altitude acclimatization. J Appl Physiol 89: 11891197
20. Gutman I and Wahlefeld WW (1974). -Lactate determination with lactate dehydrogenase and NAD. In: Methods of Enzymatic Analysis. edited by Hu B. New York: Academic 14641468
21. Hagberg J and Coyle E (1983). Physiological determinants of endurance performance as studied in competitive racewalkers. Med Sci Sports Exerc 15: 287289
22. Hansen E, Andersen J, Nielsen J, and Sjogaard G (2002). Muscle fibre type, efficiency, and mechanical optima affect freely chosen pedal rate during cycling. Acta Physiol Scand 176: 185194
23. Hickson R, Hagberg J, Ehsani A, and Holloszy J (1981). Time course of the adaptive responses of aerobic power and heart rate to training. Med Sci Sports Exerc 13: 1720
24. Hintz C, Coyle E, Kaiser K, Chi M, and Lowry O (1984). Comparison of muscle fiber typing by quantitative enzyme assays and by myosin ATPase staining. J Histochem Cytochem 32: 655660
25. Horowitz J, Sidossis L, and Coyle E (1994). High efficiency of type I muscle fibers improves performance. Int J Sports Med 15: 152157
26. Kushmerick MJ (1983). Energetics of muscle contraction. In: Handbook of Physiology. Skeletal Muscle. Bethesda, MD. MD: Am Physiol Soc, sect. 10, chapt. 7, p. 189236
27. Neptune R and Hull M. A (1999). Theoretical analysis of preferred pedaling rate selection in endurance cycling. J Biomech 32: 409415
28. Padilla S, Mujika I, Angulo F, and Goiriena JJ (2000). Scientific approach to the 1-h cycling world record: a case study. J Appl Physiol 89: 15221527
29. Pollock ML (1977). Submaximal and maximal working capacity of elite distance runners. Part I. Cardiorespiratory aspects. Ann NY Acad Sci 301: 310322
30. Saunders PU, Telford RD, Pyne DB, Cunningham RB, Gore CJ, Hahn AG, and Hawley JA (2004). Improved running economy in elite runners after 20 days of simulated moderate-altitude exposure. J Appl Physiol 96: 931937
31. Sidossis L, Horowitz J, and Coyle E (1992). Load and velocity of contraction influence gross and delta mechanical efficiency. Int J Sports Med 13: 407411
32. Widrick JJ, Trappe SW, Blaser CA, Costill DL, and Fitts RH (1996). Isometric force and maximal shortening velocity of single muscle fibers from elite master runners. Am J Physiol Cell Physiol 271: C666C675
33. Widrick JJ, Trappe SW, Costill DL, and Fitts RH (1996). Force-velocity and force-power properties of single muscle fibers from elite master runners and sedentary men. Am J Physiol Cell Physiol 271: C676C683
34. Wilmore JH (1969). A simplified method for determination of residual lung volumes. J Appl Physiol 27: 96100
35. Wilmore JH and Behnke AR (1969). An anthropometric estimation of body density and lean body weight in young men. J Appl Physiol 27: 2531
Cita Original
Coyle E.F. Improved muscular efficiency displayed as Tour de France champion matures. J. Appl. Physiol; 98: 2191-2196, 2005.
Cita en PubliCE
Edward F Coyle (2005). Mejora de la Eficiencia Muscular Desarrollada a Medida que Madura un Campeón del Tour de Francia. .https://g-se.com/mejora-de-la-eficiencia-muscular-desarrollada-a-medida-que-madura-un-campeon-del-tour-de-francia-913-sa-c57cfb2719c735