El Entrenamiento de la Fuerza No Induce Adaptaciones Uniformes al Cuádriceps
Gerald T. Mangine, Michael J. Redd, Adam M. Gonzalez, Jeremy R. Townsend, Adam J. Wells, Adam R. Jajtner, Kyle S. Beyer, Carleigh H. Boone, Michael B. La Monica, Jeffrey R. Stout, David H. Fukuda, Nicholas A. Ratamess, Jay R. HoffmanResumen
El entrenamiento de la fuerza puede afectar de forma diferente a las adaptaciones morfológicas a lo largo de la longitud de los músculos uniarticulares y biarticulares. El propósito de este estudio fue comparar los cambios en la morfología muscular a lo largo de la longitud del músculo recto femoral (RF) y el vasto lateral (VL) en respuesta al entrenamiento de la fuerza. Después de una fase preparatoria de 2 semanas, 15 hombres entrenados en fuerza (24,0 ± 3,0 años, 90,0 ± 13,8 kg, 174,9 ± 20,7 cm) completaron las evaluaciones de pre-entrenamiento (PRE) de grosor muscular (MT), ángulo de penación (PA), área de sección transversal (CSA) y ecointensidad en el RF y el VL al 30, 50 y 70% de la longitud de cada músculo; la longitud del fascículo (FL) fue estimada a partir de las mediciones respectivas del MT y el PA dentro de cada músculo y región. Luego, los participantes comenzaron un programa de entrenamiento de fuerza de alta intensidad y bajo volumen (4 x 3-5 repeticiones, 3 minutos de descanso), en miembros inferiores, y repitieron todas las PRE-evaluaciones después de 8 semanas (2 d ∙ sem-1) de entrenamiento (POST). Aunque el análisis de varianza de medidas repetidas de tres vías (músculo [RF, VL] x región [30, 50, 70%] x tiempo [PRE, POST]) no reveló interacciones significativas para ninguna evaluación de la morfología, se observaron efectos simples significativos (músculo x tiempo) para el CSA (p = 0,002) y la FL (p = 0,016). Específicamente, los cambios promedio del CSA favorecieron al VL (2,96 ± 0,69 cm2, p < 0,001) sobre el RF (0,59 ± 0,20 cm2, p = 0,011), mientras que se observaron disminuciones significativas en la FL promedio para el RF (-1,03 ± 0,30 cm, p = 0,004) pero no para el VL (-0,05 ± 0,36 cm, p = 0,901). No se observaron otras diferencias significativas. Los resultados de este estudio demuestran la aparición de adaptaciones no homogéneas en el tamaño y la arquitectura de los músculos RF y VL después de 8 semanas de entrenamiento de fuerza de alta intensidad en hombres entrenados en fuerza. Sin embargo, el entrenamiento no parece influir en las adaptaciones específicas de cada región en ninguno de los dos músculos.
Palabras Clave: fuerza, adaptaciones, cuádriceps
Abstract
Resistance training may differentially affect morphological adaptations along the length of uni-articular and bi-articular muscles. The purpose of this study was to compare changes in muscle morphology along the length of the rectus femoris (RF) and vastus lateralis (VL) in response to resistance training. Following a 2-wk preparatory phase, 15 resistance-trained men (24.0 ± 3.0 y, 90.0 ± 13.8 kg, 174.9 ± 20.7 cm) completed pre-training (PRE) assessments of muscle thickness (MT), pennation angle (PA), cross-sectional area (CSA), and echo-intensity in the RF and VL at 30, 50, and 70% of each muscle's length; fascicle length (FL) was estimated from respective measurements of MT and PA within each muscle and region. Participants then began a high intensity, low volume (4 x 3-5 repetitions, 3min rest) lower-body resistance training program, and repeated all PRE-assessments after 8 weeks (2 d ∙ wk-1) of training (POST). Although three-way (muscle [RF, VL] x region [30, 50, 70%] x time [PRE, POST]) repeated measures analysis of variance did not reveal significant interactions for any assessment of morphology, significant simple (muscle x time) effects were observed for CSA (p = 0.002) and FL (p = 0.016). Specifically, average CSA changes favored the VL (2.96 ± 0.69 cm2, p < 0.001) over the RF (0.59 ± 0.20 cm2, p = 0.011), while significant decreases in average FL were noted for the RF (-1.03 ± 0.30 cm, p = 0.004) but not the VL (-0.05 ± 0.36 cm, p = 0.901). No other significant differences were observed. The findings of this study demonstrate the occurrence of non-homogenous adaptations in RF and VL muscle size and architecture following 8 weeks of high-intensity resistance training in resistance-trained men. However, training does not appear to influence region-specific adaptations in either muscle.
Keywords: strength, adaptations, quadriceps
INTRODUCCIÓN
La selección y modalidad del ejercicio influyen en el grado de activación de músculos específicos durante el entrenamiento. La activación aumenta cuando los ejercicios se vuelven más complejos [1], mientras que el rango de movimiento puede alterar la contribución porcentual de varios grupos musculares asociados con el ejercicio [2]. Estas diferencias parecen estar moduladas por el papel específico de cada músculo durante el movimiento. Por ejemplo, el músculo biarticular recto femoral (RF) y el músculo monoarticular vasto lateral (VL) poseen una función similar durante la extensión de rodilla, y por lo tanto, se activan de manera similar durante ese ejercicio [3]. Sin embargo, sus roles funcionales son diferentes cuando un ejercicio requiere movimiento simultáneo en las articulaciones de cadera y rodilla (por ejemplo, sentadilla o peso muerto) [4]. Durante la fase de descenso en sentadilla o peso muerto las fibras proximales del RF se acortan para flexionar la cadera, mientras que las fibras distales del RF y del VL se alargan para flexionar la rodilla. Este proceso se invierte durante el ascenso. Dado que la intensidad relativa (es decir, el porcentaje de la fuerza máxima) variará a lo largo del movimiento dinámico, debido a los cambios en la velocidad y la ventaja mecánica [4,5], es posible que el grado de exposición al estímulo también sea diferente entre el RF y el VL. De hecho, se ha observado que la activación del RF es un 32% mayor durante la fase de ascenso de una sentadilla en comparación con la de descenso, mientras que la contribución del VL se mantiene constante [6]. La producción de fuerza relativa también parece ser diferente entre el RF y el VL durante la sentadilla [7], aunque esto no se ha evaluado estadísticamente. En consecuencia, estas diferencias agudas pueden afectar a las adaptaciones del entrenamiento.
Las adaptaciones al músculo esquelético se consideran específicas a la demanda impuesta del ejercicio [5,8] con cambios en su composición metabólica y estructural que reflejan los requisitos funcionales [9]. Las fibras musculares esqueléticas activadas se hipertrofiarán en respuesta al estrés mecánico y a la fatiga inducida por sesiones de entrenamiento repetidas [10,11]. Sin embargo, no se puede esperar un crecimiento uniforme en cada músculo. Se ha observado que los cambios en la arquitectura muscular varían entre los músculos [12, 13], así como el grosor [14,15] y la longitud [12,13] de músculos específicos. Estas diferencias parecen estar afectadas por la modalidad del entrenamiento y, potencialmente, por la experiencia en el entrenamiento. En hombres no entrenados, Narici y colegas (1996) informaron diferencias en la hipertrofia entre cada uno de los músculos del cuádriceps después de 6 meses de extensiones unilaterales de pierna realizadas en días alternos, y que los cambios favorecieron las porciones más distales del RF y el VL. Asimismo, se ha observado que las adiciones (o pérdidas) de sarcómeros en serie se producen a lo largo del grosor y profundidad del tibial anterior después de 6 semanas de entrenamiento excéntrico utilizando varias posiciones de partida (es decir, el grado de flexión plantar) en conejos [15]. En contraste, se ha documentado una mayor hipertrofia del VL (regiones medias a distales) en comparación con la hipertrofia limitada del RF en mujeres mayores no entrenadas cuando el entrenamiento incluía tanto un ejercicio monoarticular como uno multiarticular (es decir, prensa de piernas) [13]. Aunque estos hallazgos destacan la aparición de adaptaciones no homogéneas en todo el músculo esquelético, también se han documentado cambios uniformes siguiendo un protocolo de entrenamiento similar (es decir, 5 semanas de extensiones de pierna) [16]. Estos hallazgos pueden estar limitados por la experiencia en entrenamiento de los participantes y la simplicidad del programa respectivo de cada estudio. Se observaron mayores adaptaciones mediales (5 cm desde la línea media) en el grosor del VL (y posiblemente en el ángulo de penación) en comparación con las encontradas en la línea media, después de un protocolo de 15 semanas, periodizado, de método mixto (es decir, entrenamiento de fuerza, olímpico y pliométrico) [14]. Aún así, el programa estaba destinado a desarrollar la fuerza y la potencia para el rendimiento deportivo en los atletas de fútbol de la División I; la hipertrofia muscular era un objetivo secundario de entrenamiento. Por lo tanto, aún no está claro si los cambios en la arquitectura muscular serían homogéneos después de un protocolo diseñado para el crecimiento muscular en individuos entrenados.
Cuando se entrena para la hipertrofia, se espera que se añadan proteínas contráctiles a los sarcómeros existentes [17] para aumentar el diámetro y la longitud de la fibra, haciéndola más fuerte y duradera contra el daño futuro causado por el mismo estímulo [18]. Este efecto es más pronunciado en los levantadores de pesas no entrenados porque la mayoría de los diseños de entrenamiento son novedosos para esta población y sus fibras musculares aún no han desarrollado una "resistencia" a varios estímulos. Por lo tanto, es posible que las pequeñas diferencias en el programa no alteren la respuesta del entrenamiento. Por ejemplo, la suma de ejercicios monoarticulares (es decir, extensión de tríceps y flexión de codo) a un programa de entrenamiento de fuerza multiarticular no resultó en una mayor hipertrofia para hombres no entrenados [19]. Por el contrario, se necesita mayor precisión en las características del programa (es decir, intensidad, volumen, densidad) para iniciar este proceso en adultos entrenados [5,8]. Para estos individuos, se ha encontrado que la frecuencia de la serie [20], la intensidad y el volumen del entrenamiento [21,22] y los intervalos de descanso [23] influyen en la respuesta a la hipertrofia. Sin embargo, en varios casos, la hipertrofia observada no fue consistente en cada sitio [21-23], ni se compararon estas diferencias. Por lo tanto, el propósito de este estudio fue comparar los cambios entre la arquitectura del RF y el VL a lo largo de su eje longitudinal después de 8 semanas de entrenamiento de fuerza en hombres entrenados en fuerza. Basándonos en informes anteriores [12-14,24], planteamos la hipótesis de que los cambios en la arquitectura muscular serían diferentes entre el RF y el VL, y que estos cambios variarían a lo largo de la longitud de cada músculo.
MATERIALES Y MÉTODOS
Diseño del estudio
Las reducciones en las características del músculo esquelético (es decir, el tamaño y la arquitectura) pueden ocurrir dentro de las dos semanas posteriores al desentrenamiento (es decir, el cese del entrenamiento) en poblaciones entrenadas en fuerza [25]. Por lo tanto, para evaluar el efecto del entrenamiento sobre las adaptaciones musculares en las regiones del RF y el VL, este estudio no empleó el uso de un grupo control. En cambio, se utilizó un diseño dentro de los sujetos en el que se compararon las evaluaciones de la morfología muscular pre-entrenamiento (PRE) con las observadas después de 8 semanas de entrenamiento de fuerza (POST). Inicialmente, todos los participantes se presentaron al Laboratorio de Desempeño Humano (HPL) para completar un programa obligatorio de entrenamiento preparatorio de 2 semanas. Posteriormente, se realizaron PRE-evaluaciones de la morfología muscular en todos los participantes. Los participantes regresaron al HPL la semana siguiente (es decir, la semana 3) para comenzar el programa de entrenamiento de 8 semanas. Durante la semana siguiente a la intervención del entrenamiento de 8 semanas (es decir, la semana 11), se repitieron todas las PRE-evaluaciones. Se hicieron comparaciones entre músculos y entre regiones a lo largo del tiempo.
Participantes
Después de una explicación de todos los procedimientos, riesgos y beneficios, 15 hombres físicamente activos y entrenados en fuerza (24,0 ± 3,0 años; 90,0 ± 13,8 kg; 174,9 ± 20,7 cm) dieron su consentimiento informado por escrito para participar en este estudio. Todos los participantes estaban libres de cualquier limitación física (determinada por el cuestionario de historia clínica y el PAR-Q) y habían estado participando regularmente en el entrenamiento de fuerza durante un mínimo de 2 años (5,7 ± 2,2 años) en el momento del reclutamiento. Los participantes describieron que sus hábitos de entrenamiento previos eran diferentes del régimen de entrenamiento actual en términos de orden de ejercicio y agrupaciones. Aproximadamente el 87% describió su rango de repetición normal como más alto (es decir, rango de 6-12 RM) que el rango de 3-5 RM usado en este estudio. Además, el 47% informó haber usado períodos de descanso más cortos (es decir, < 3 minutos), mientras que el 13,3% no había hecho un seguimiento de sus tiempos de descanso anteriormente. El resto de los participantes emplearon un esquema de entrenamiento similar al del programa utilizado en este estudio. Esta investigación fue aprobada por la Junta de Revisión Institucional de New England.
Mediciones por Ecografía
Después de 15 minutos de reposo en posición supina, para permitir la redistribución de los fluidos corporales [26], se tomaron imágenes ecográficas del RF y el VL del muslo dominante de cada participante utilizando un cabezal de exploración de sonda lineal de 12 MHz (General Electric LOGIQ P5, Wauwatosa, WI, EEUU). El mismo investigador identificó todos los lugares anatómicos de interés utilizando los estándares de referencia descritos anteriormente [26-28] para medir el grosor muscular (MT; ±0,1 cm), el CSA (±0,1 cm2), la ecointensidad (EI; ±0,1 unidades arbitrarias [au]) y el ángulo de penación (PA; ±0,1°). Para cada músculo, se recolectaron imágenes de distal a proximal a lo largo de la distancia longitudinal de la línea media al 30% (es decir, distal), 50% (es decir, medio) y 70% (es decir, proximal) de la longitud de cada músculo. Para el CSA y la EI, el modo de campo visual extendido (Ganancia = 50dB; Profundidad = 5cm) se utilizó para capturar dos imágenes panorámicas consecutivas de las regiones musculares de interés. Para el MT y el PA, se recolectaron dos imágenes de los mismos sitios descritos para el CSA y la EI, pero con la sonda orientada longitudinalmente a la interfase del tejido muscular usando ecografía Modo Luminosidad (modo-B). Todas las imágenes recolectadas fueron transferidas a una computadora personal y analizadas por el mismo investigador usando la Imagen J (National Institutes of Health, Bethesda, MD, EEUU, versión 1.45s).
Los valores promediados de ambas imágenes para cada medida dentro de una región específica se utilizaron para el análisis estadístico. La longitud del fascículo (FL; ± 0,1 cm) para cada músculo dentro de cada región se estimó usando imágenes asociadas para el MT y el PA. Esta metodología para la determinación de la longitud del fascículo tiene un coeficiente de variación estimado informado de 4,7% [29] y se puede encontrar usando la siguiente ecuación [29-31]: Fascículo = MT • SIN (PA)-1. La fiabilidad de estos procedimientos para evaluar el MT (ICC3.K = 0,88-0,92, SEM3.K = 0,15-0,39 cm), CSA (ICC3.K = 0,88-0,99, SEM3.K = 0,81-2,38 cm2), EI (ICC3.K = 0,74-0,95, SEM3.K = 2,59-6,44 au), PA (ICC3.K = 0,81-0,97, SEM3.K = 0,27-1,44°) y FL (ICC3.K = 0,81-0,96, SEM3.K = 0,74-1,35 cm) al 30%, 50% y 70% de la longitud del RF y el VL se habían determinado previamente en 10 hombres activos y entrenados en fuerza (25,3 ± 2,0 años; 90,8 ± 6,8 kg; 180,3 ± 7,1 cm).
Intervención de Entrenamiento de Fuerza
Los detalles del entrenamiento y los tests de fuerza se han descrito en otra parte [21]. Brevemente, todos los participantes completaron una fase preparatoria de 2 semanas antes de la intervención de 8 semanas para familiarizarse con los ejercicios de entrenamiento, el protocolo y la técnica de levantamiento adecuada. El rendimiento durante esta fase, junto con la fuerza de una repetición máxima (1RM) evaluada en la sentadilla, se utilizó para calcular las cargas de intensidad utilizadas durante la intervención. El programa de entrenamiento de alta intensidad y bajo volumen (4 series de 3 a 5 repeticiones, intervalos de descanso de 3 minutos) utilizado en este estudio incluyó cuatro ejercicios de cadena cerrada de miembros inferiores (es decir, sentadilla con barra, peso muerto, estocada con barra y prensa de piernas) que se realizaron en dos sesiones de entrenamiento por semana. La carga de intensidad inicial se fijó en el 90% de 1RM evaluada (sentadilla) o estimada (todos los demás ejercicios) [32] de cada participante. Las cargas de entrenamiento se incrementaron progresivamente cuando todas las repeticiones prescritas para un ejercicio se lograron en dos entrenamientos consecutivos. Se pidió a todos los participantes que completaran al menos 14 (de 16) sesiones de entrenamiento (~87,5%). Todas las sesiones se realizaron bajo la supervisión directa de especialistas certificados en fuerza y acondicionamiento (CSCS).
Ingesta de Nutrientes y Análisis Alimentario
Durante la intervención del entrenamiento, se instruyó a los participantes para que mantuvieran sus hábitos alimenticios normales. Para asegurar que la nutrición después del ejercicio fuera consistente, cada participante recibió ~235 mL de leche chocolatada (170 calorías; 2,5 g de grasa; 29 g de carbohidratos; 9 g de proteína), o Lactaid® (150 calorías; 2,5 g de grasa; 24 g de carbohidratos; 8 g de proteína) para los participantes intolerantes a la lactosa, después de cada sesión de entrenamiento. Además, la ingesta total de kilocalorías y macronutrientes de todas las fuentes de alimentos y bebidas se controló a través de diarios alimentarios de tres días (dos días de la semana y un día del fin de semana), dado el efecto que cualquier cambio tendría en la adaptación muscular. Estos diarios fueron recogidos durante la primera y última semana de la intervención del entrenamiento. El software FoodWorks Dietary Analysis versión 13 (The Nutrition Company, Long Valley, NJ) se utilizó para analizar cada diario alimentario. Para el análisis estadístico, se analizó la ingesta total de calorías y proteínas en relación con la masa corporal.
Análisis Estadístico
Se utilizó un software estadístico (V. 24.0, SPSS Inc., Chicago, IL) para determinar si existían diferencias entre regiones y músculos después del entrenamiento. Los datos se analizaron mediante análisis de varianza de medidas repetidas de tres vías (músculo [RF, VL] x región [30%, 50%, 70%] x tiempo [PRE, POST]) (RM_ANOVA) con medidas repetidas para cada medición de morfología muscular. Las interacciones significativas entre los factores y los efectos principales simples se examinaron más a fondo mediante un RM_ANOVA bidireccional separado (es decir, región x tiempo, músculo x tiempo) y aplicando los ajustes de Bonferroni a los intervalos de confianza cuando fue apropiado. La significación estadística se fijó en un nivel alfa de p ≤ 0,05. Las diferencias observadas se evaluaron en profundidad mediante el uso de tamaños del efecto (η2P: Partial eta squared) y los siguientes niveles: efecto pequeño (0,01-0,058), efecto medio (0,059-0,137) y efecto grande (>0,138) [33]. Para evaluar si las diferencias observadas podían considerarse reales, se compararon los cambios con su diferencia mínima calculada (MD) [34] mediante la creación de un intervalo de confianza (IC) del 95% sobre el error estándar de la medición (SEM). La MD se calculó utilizando la siguiente ecuación (MD = SEM x 1,96 x √2). Cualquier cambio que ocurriera dentro de este intervalo de confianza se interpretó como consistente con el error de medición de la prueba, mientras que los cambios que ocurrieran fuera del intervalo reflejan cambios reales en la composición corporal. Todos los datos se informan como media ± error estándar (SE) de la media.
Resultados
Después de 8 semanas de entrenamiento, se observó un efecto principal significativo del tiempo para el CSA (F = 19,9; p<0,001; ɳ2P = 0,59), donde el tamaño medio de los músculos (es decir, la combinación de RF y VL) aumentó en 1,78 ± 0,40 cm2 (IC del 95% = 0,92-2,63 cm2). Además, se observaron tendencias para el tiempo en las que el promedio del PA aumentó (F = 4,1, p=0,063, ɳ2P = 0,23) en 0,69 ± 0,34° (IC del 95% = -0,04-1,43) y el promedio de la FL disminuyó (F = 3,7, p=0,076, ɳ2P = 0,21) en -0,54 ± 0,28 (IC del 95% = -1,14-0,07). En la Tabla 1 se presentan las estimaciones marginales para las medidas de la morfología muscular a lo largo de 8 semanas de entrenamiento.
Tabla 1. Cambios en las estimaciones marginales de la morfología combinada del RF y el VL tras 8 semanas de entrenamiento de fuerza (media ± SE).
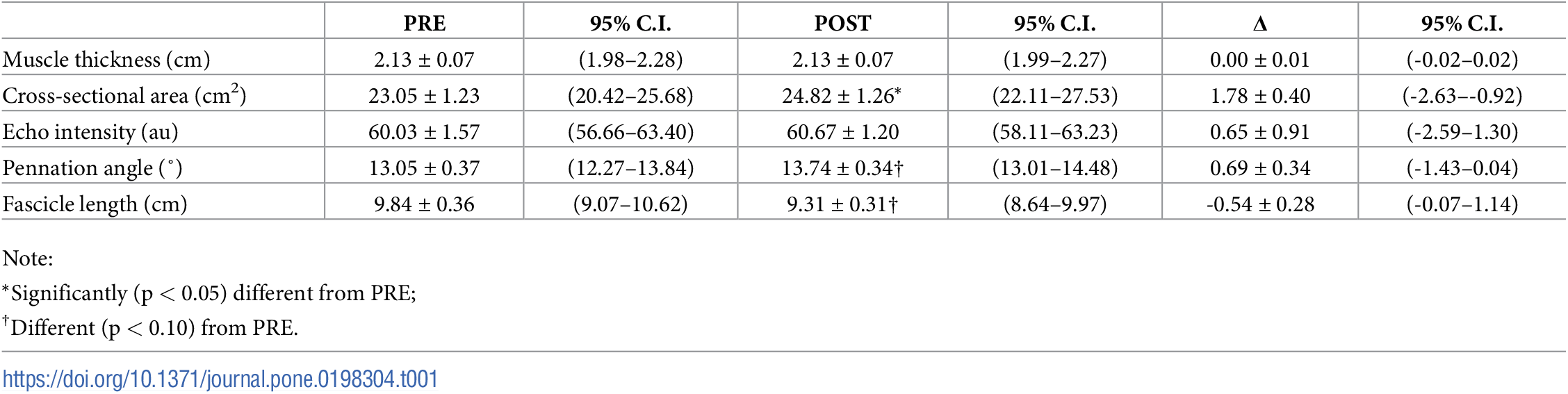
El ANOVA de tres vías no reveló una interacción músculo x región x tiempo significativa para el MT (F = 0,3; p = 0,741; ɳ2P = 0,02), el CSA (F = 1,9; p = 0,189; ɳ2P = 0,12), el PA (F = 0,3, p = 0,757, ɳ2P = 0,02), o la FL (F = 0,6, p = 0,544, ɳ2P = 0,04), aunque se observó una tendencia para la EI (F = 3,2, p = 0,057, ɳ2P = 0,19). El análisis exploratorio post-hoc reveló un aumento significativo de la EI del RF al 70% (3,70 ± 1,09 au, p = 0,004), pero no en ningún otro lugar.
Se observó un efecto simple (músculo x tiempo) significativo para el CSA (F = 14,1, p = 0,002, ɳ2P = 0,50) donde los cambios favorecieron al VL (2,96 ± 0,69 cm2, IC del 95% = 1,48-4,44 cm2, p < 0,001) sobre el RF (0,59 ± 0,20 cm2, IC del 95% = 0,15-1,03 cm2, p = 0,011). También se observó un efecto simple (músculo x tiempo) significativo para la FL (F = 7,5, p = 0,016, ɳ2P = 0,35) donde el RF disminuyó (-1,03 ± 0,30 cm, IC del 95% = -1,68 –0,38 cm, p = 0,004) y el VL no cambió (-0,05 ± 0,36 cm, IC del 95% = -0,82-0,73 cm, p = 0,901). Además, se observaron tendencias para el MT (F = 4,3, p = 0,058, ɳ2P = 0,23) y el PA (F = 3,3, p = 0,091, ɳ2P = 0,19). Las diferencias entre músculos y regiones en PRE y POST para cada evaluación de la morfología muscular se ilustran en la Fig. 1.
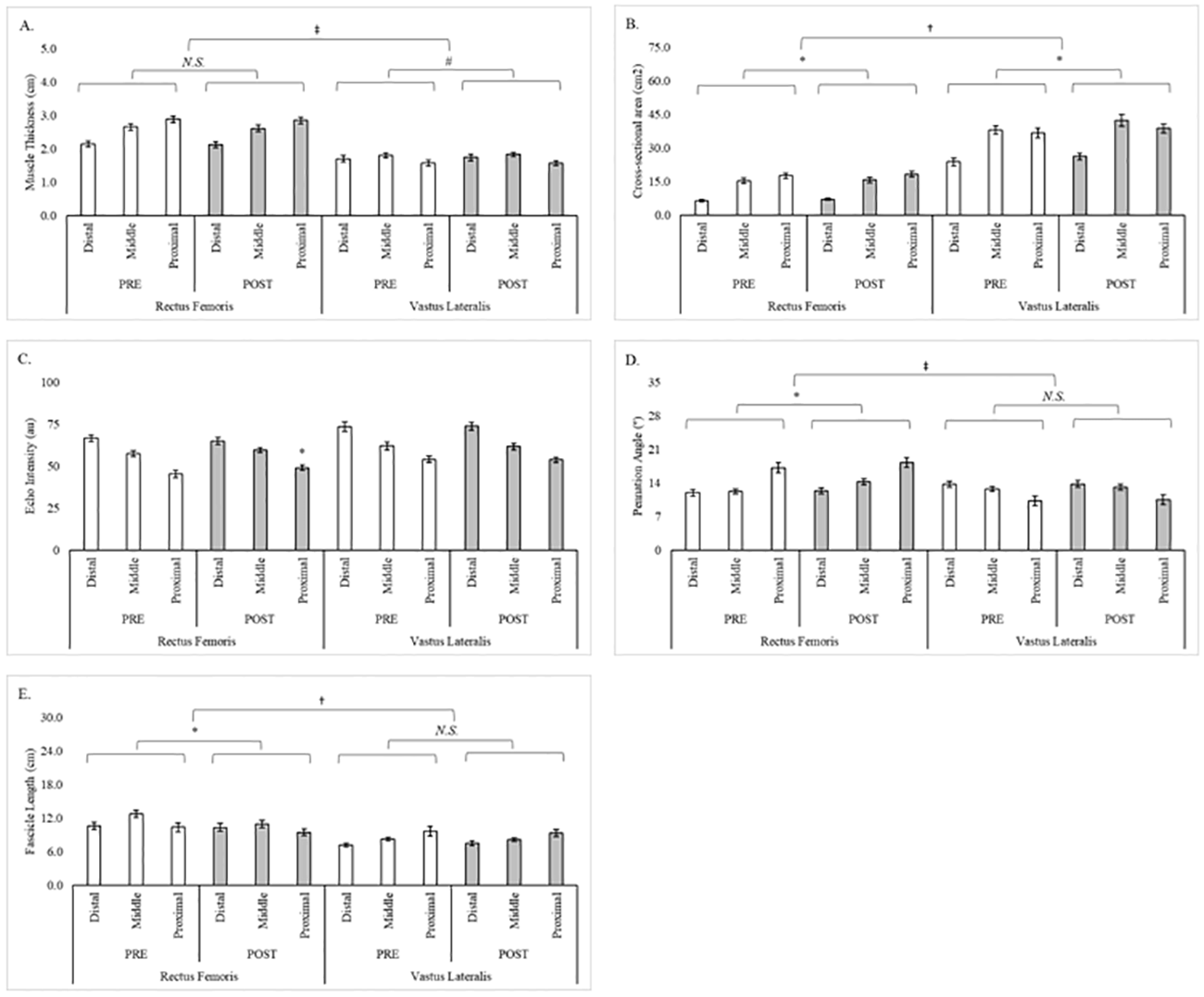
Figura 1. Diferencias regionales y musculares en la morfología muscular a lo largo de 8 semanas de entrenamiento de fuerza. (A. Grosor del músculo; B. Área de sección transversal; C. Ecointensidad; D. Ángulo de penación; y E. Longitud del fascículo).
Nota: *= Diferencia significativa (p < 0,05) entre PRE y POST; †= Diferencias significativas (p < 0,05) entre RF y VL; # = Diferencia (p < 0,10) entre PRE y POST; ‡ = Diferencia (p < 0,10) entre RF y VL; N.S. = No significativa.
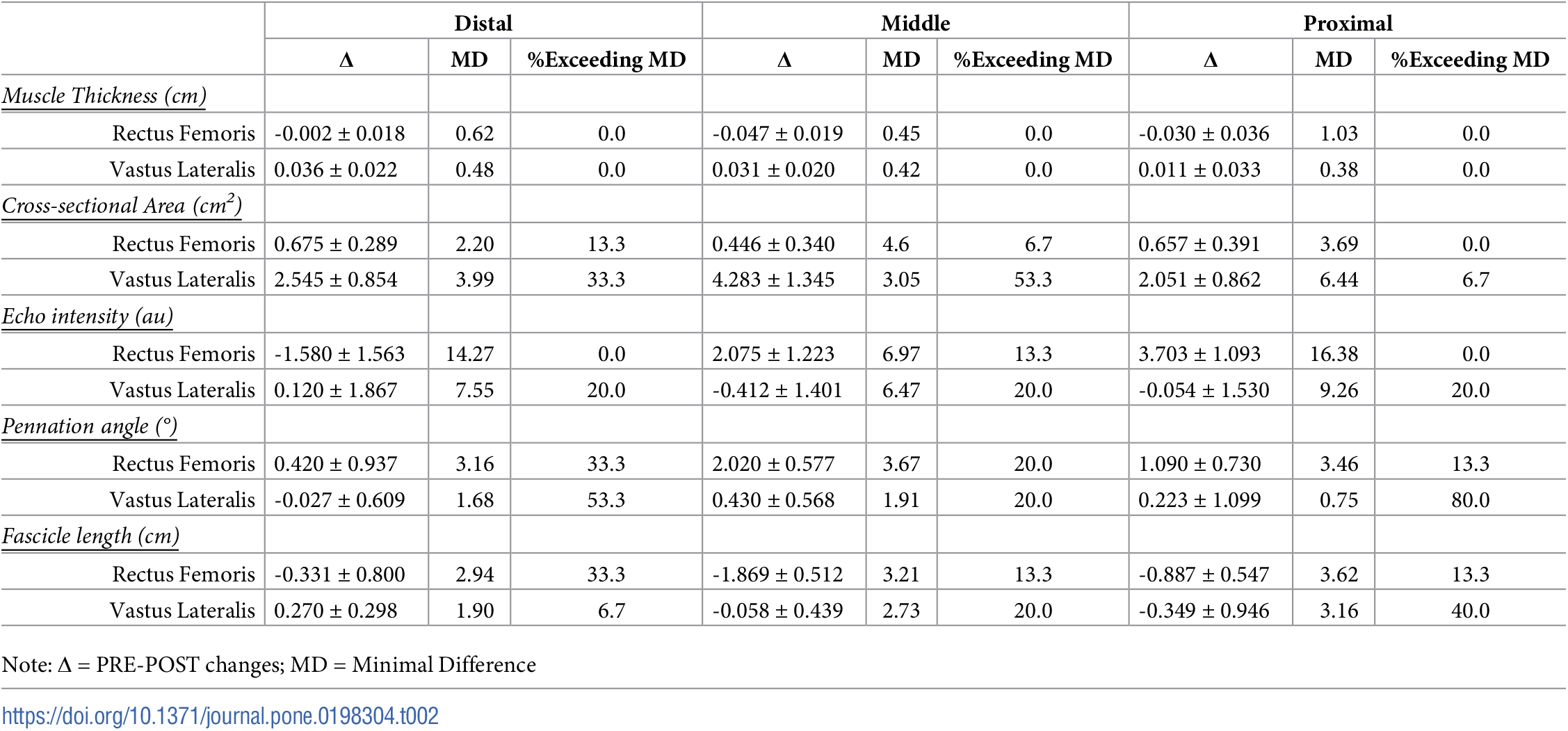
Aunque no se observaron otras diferencias estadísticas, un mayor porcentaje de participantes experimentó cambios en la morfología del VL en comparación con el RF para el CSA (todas las regiones), la EI (todas las regiones), el PA (30% y 70%) y la FL (50% y 70%). Se observaron adaptaciones similares entre los músculos para la el PA al 50% (20% para RF y VL), mientras que un mayor porcentaje de participantes superó la MD para la FL al 30% en el RF (33,3%) en comparación con el VL (6,7%). Dentro del RF, un mayor porcentaje de participantes experimentó cambios que excedieron la MD en un 30% (CSA, PA y FL) en comparación con otras regiones. En contraste, los cambios que excedían la MD para cada región del VL variaron según la evaluación morfológica. Los cambios en el MT (RF y VL) no superaron el MD de ninguna región. En la Tabla 2 se presentan los cambios regionales en la morfología del RF y el VL y el porcentaje de participantes que superan la MD respectiva de cada medida.
Tabla 2. Porcentaje de participantes que superan la diferencia mínima para cambios regionales en la morfología muscular después de 8 semanas de entrenamiento de fuerza (media ± SE).
No se observaron diferencias en la ingesta relativa de kilocalorías o proteínas a lo largo de 8 semanas de entrenamiento de fuerza ni se han informado previamente en otro lugar [21].
Discusión
El propósito de esta investigación fue determinar si los cambios en la morfología del RF y el VL fueron uniformes después de 8 semanas de entrenamiento de fuerza en hombres entrenados en fuerza. Se planteó la hipótesis de que los cambios diferirían entre estos músculos, así como a lo largo de sus ejes longitudinales, debido a las diferencias en su rol funcional durante los ejercicios multiarticulares de los miembros inferiores (p.ej., sentadilla y peso muerto) [4]. En general, observamos una mayor hipertrofia en el VL en comparación con el RF, lo que fue consistente con lo observado por Häkkinen y colegas (2001) pero no con otros [12,16,24]. Después del entrenamiento que sólo incluyó ejercicios de cadena abierta (es decir, extensiones de pierna), se observó una hipertrofia del RF similar [16] o mayor [12,24], en comparación con el VL. Además, observamos una disminución de la FL y una tendencia a un aumento del PA en el RF sin que se produjeran cambios en el VL. Estos hallazgos difirieron de los de Seynnes y colegas (2007) que informaron aumentos uniformes en la FL y el PA. Además, aparte de una tendencia hacia la disminución de la calidad muscular del RF proximal (vía EI), no se encontraron diferencias regionales específicas en las adaptaciones morfológicas. Anteriormente, se habían informado diferencias a lo largo del eje longitudinal [12,13,24] después de protocolos de entrenamiento de fuerza que sólo incluían extensiones unilaterales de pierna [12,24] o sólo incluían un único ejercicio multiarticular [13] en adultos con experiencia limitada de entrenamiento. Sin embargo, nuestro estudio parece ser el primero en examinar las diferencias regionales en los cambios musculares después de un programa de entrenamiento que sólo utilizó ejercicios multiarticulares en un grupo de hombres entrenados en fuerza.
El grupo muscular del cuádriceps está compuesto por cuatro músculos que se insertan en el tendón rotuliano pero que se originan en varias estructuras de la cadera y el fémur. Debido a las diferencias de origen, sus funciones individuales se ven afectadas por el movimiento. Durante la fase concéntrica de una extensión de pierna, los músculos del cuádriceps se activan juntos para contribuir igualmente a la producción de fuerza [35,36]; aunque la activación del RF puede aumentar durante la fase excéntrica [12]. Por el contrario, la activación del RF es menor que la del VL durante un ejercicio de cadena cerrada (por ejemplo, prensa de piernas, sentadilla) [35-37] y también puede ser menor durante la fase excéntrica en comparación con la fase concéntrica [6]. Anteriormente, se ha encontrado una mayor hipertrofia del RF (que del VL) cuando el entrenamiento sólo incluía extensiones de pierna [12,24]. Sin embargo, nuestro protocolo de entrenamiento sólo incluyó ejercicios de cadena cerrada y resultó en una mayor hipertrofia del VL. Cuando el entrenamiento ha incluido previamente un ejercicio de cadena cerrada, las adaptaciones han favorecido al VL [13]. Por lo tanto, es posible que las mayores adaptaciones del VL, y la posible reducción de la calidad del RF, se hayan relacionado con que nuestro diseño del programa sea más específico para la activación del VL. Aunque esto no puede ser confirmado por nuestro informe anterior de cambios similares en la activación del VL (-1,62 ± 1,80 V ∙ seg ∙ %1RM-1) y del RF (-1,72 ± 1,47 V ∙ seg ∙ %1RM-1) a través de pruebas de fuerza máxima y submáxima [21], la activación sólo se evaluó al 50% de la longitud del músculo. No está claro si existen diferencias de adaptación en la activación a lo largo de cada músculo después de este tipo de programa en individuos entrenados.
Además de las diferencias en la hipertrofia, también se observaron diferencias en los cambios de la arquitectura entre los músculos. Se produjo un aumento del PA y una disminución de la FL en el RF, mientras que no se observaron cambios en el VL. Se cree que los cambios en el tamaño muscular afectan a la arquitectura muscular [14,24,38-40]. Específicamente, el aumento del MT se ha asociado con un aumento del PA [24,38,39] pero no de la FL [14, 24, 39], aunque se ha descubierto que los individuos que poseen una mayor CSA tienen un mayor PA y una mayor FL [40]. Después del ejercicio de fuerza, las áreas dañadas del músculo están habitadas por células satélites, que se fusionan con el tejido muscular existente [41] y añaden nuevas proteínas contráctiles que aumentan el diámetro de los sarcómeros existentes y la longitud de las fibras [17]. Aunque los cambios en el grosor del músculo y el diámetro de la fibra deben afectar la orientación de la fibra y el ángulo de inserción, los cambios en la FL pueden depender de la modalidad del ejercicio. Cuando el entrenamiento se compone predominantemente de acciones de alargamiento muscular (es decir, entrenamiento excéntrico), se agrega un mayor número de sarcómeros en forma serial en comparación con las contracciones concéntricas o mixtas [42,43]. Aquí, el protocolo de entrenamiento incluía ejercicios que incorporaban tanto contracciones excéntricas como concéntricas. Sin embargo, el grado y la duración de la tensión excéntrica pueden haber variado en función de la técnica individual (por ejemplo, la velocidad de descenso de la barra durante el peso muerto, el grado de implicación del extensor de cadera, el rango de movimiento). Esta variabilidad se reflejó en que los errores estándar para los cambios en la FL fueron mayores que sus respectivas medias, así como en el porcentaje de participantes que excedieron la MD necesaria para observar los cambios "reales" en cada lugar de medición (ver Tabla 2). También es posible que no se hayan realizado las adaptaciones de la FL debido a que las mediciones utilizadas para la estimación de la FL se recogieron a lo largo de la línea media de cada músculo. Anteriormente, Wells y colegas (2014) informaron diferencias entre los cambios en la FL observados a lo largo de la línea media del VL (a un 50% de la longitud del músculo) y un sitio ubicado a 5 cm medialmente. Es posible que nuestro protocolo de entrenamiento, la forma en que los participantes realizaron los ejercicios y los sitios específicos utilizados para la estimación de la FL, limitaron nuestro potencial para observar mejoras en la FL.
Aparte de una tendencia hacia un aumento de la EI (en el RF proximal), nuestros datos no apoyaron la hipótesis de que las adaptaciones diferirían entre las regiones musculares. Anteriormente, se ha informado de que los cambios morfológicos a lo largo del eje longitudinal del RF y el VL son equívocos [12,13,16,24] cuando los ejercicios multiarticulares se utilizan escasamente o son inexistentes en adultos con experiencia en entrenamiento limitada. Cuando el programa sólo incluía extensiones de piernas, se encontró que la hipertrofia era mayor en las regiones distales del cuádriceps después de 3-6 meses de entrenamiento [12,24]. Por el contrario, Häkkinen y colegas (2001) observaron hipertrofia similar a lo largo del cuádriceps, pero no cuando se consideraron los músculos individuales. Curiosamente, Seynnes y colegas (2007) no informaron diferencias entre la región distal y el vientre muscular. Sin embargo, esos hallazgos pueden haber sido limitados por un período de entrenamiento mucho más corto (por ejemplo, 5 semanas) y un análisis estadístico inapropiado (por ejemplo, t-Tests apareados por separado). Como hemos discutido anteriormente, la falta de diferencias regionales puede estar relacionada con el reclutamiento del cuádriceps durante varias modalidades de ejercicio. La activación del cuádriceps favorece las regiones distales durante una extensión de pierna de cadena abierta, mientras que una mayor activación proximal ocurre en tareas que requieren flexión activa de cadera [44]. Durante los movimientos complejos (por ejemplo, al caminar) se ha demostrado que la contribución de las regiones proximales y distales del RF varían a lo largo del movimiento y en relación con la velocidad [45]. Sin embargo, más allá de estos estudios, se sabe poco acerca de las diferencias regionales en la activación del cuádriceps durante los ejercicios multiarticulares de cadena cerrada. Es posible que la tendencia observada en la reducción de la calidad del RF proximal (es decir, el aumento de la EI) pueda ser indicativa de un efecto de desentrenamiento provocado por una reducción de la contribución de esta región a lo largo del entrenamiento.
Los resultados de este estudio demuestran la existencia de adaptaciones no homogéneas en la morfología del RF y el VL tras 8 semanas de entrenamiento de fuerza en hombres entrenados en fuerza. El programa de entrenamiento resultó en una mayor hipertrofia del VL, que puede haber sido la consecuencia de la reducción de la contribución del RF durante los ejercicios de cadena cerrada, multiarticulares. Además, el alto grado de variabilidad en el que se pueden realizar estos ejercicios (por ejemplo, la velocidad de descenso de la barra durante el peso muerto, el grado de compromiso del extensor de cadera, el rango de movimiento) puede haber sido responsable del aumento observado en el PA y la disminución en la FL del RF. Sin embargo, contrariamente a nuestra hipótesis, no observamos diferencias entre regiones (es decir, proximal, media y distal) de ninguno de los dos músculos, salvo una tendencia en la reducción de la calidad del RF proximal. Debido a que se sabe poco acerca de la activación del cuádriceps específica de la región durante los ejercicios de cadena cerrada multiarticulares, aún no está claro por qué las adaptaciones regionales fueron uniformes. Sin embargo, puede ser aconsejable que los entrenadores de fuerza y los atletas incorporen ejercicios de flexión de cadera dentro de los programas de entrenamiento de fuerza de miembros inferiores para evitar posibles reducciones en la calidad del RF proximal.
Referencias
1. Boudreau SN, Dwyer MK, Mattacola CG, Lattermann C, Uhl TL, McKeon JM. (2009). Hip-muscle activation during the lunge, single-leg squat, and step-up-and-over exercises. Journal of Sport Rehabilitation. 2009;18(1):91–103. pmid:19321909
2. Caterisano A, Moss RE, Pellinger TK, Woodruff K, Lewis VC, Booth W, et al. (2002). The effect of back squat depth on the EMG activity of 4 superficial hip and thigh muscles. J Strength Cond Res. 2002;16(3):428–32. pmid:12173958
1. Purkayastha S, Cramer JT, Trowbridge CA, Fincher AL, Marek SM. (2006). Surface electromyographic amplitude-to-work ratios during isokinetic and isotonic muscle actions. Journal of athletic training. 2006;41(3):314. pmid:17043700
2. Floyd RT. (2014). Manual of Structural Kinesiology.
3. Haff GG, Triplett NT. (2015). Essentials of Strength Training and Conditioning 4th Edition: Human kinetics.
4. McCaw ST, Melrose DR. (1999). Stance width and bar load effects on leg muscle activity during the parallel squat. Med Sci Sports Exerc. 1999;31:428–36. pmid:10188748
5. Escamilla RF, Fleisig GS, Zheng N, Lander JE, Barrentine SW, Andrews JR, et al. (2001). Effects of technique variations on knee biomechanics during the squat and leg press. Med Sci Sports Exerc. 2001;33(9):1552–66. pmid:11528346
6. Ratamess NA, Alvar BA, Evetoch TK, Housh TJ, Kibler WB, K WJ, et al. (2009). Progression models in resistance training for healthy adults. Position Stand: Medicine and science in sports and exercise. 2009;41(3):687–708. pmid:19204579
7. Flück M. (2006). Functional, structural and molecular plasticity of mammalian skeletal muscle in response to exercise stimuli. J Exp Biol. 2006;209(12):2239–48.
8. Moritani T. (1993). Neuromuscular adaptations during the acquisition of muscle strength, power and motor tasks. J Biomech. 1993;26:95–107.
9. Evans WJ. (2002). Effects of exercise on senescent muscle. Clin Orthop Relat Res. 2002;403:S211–S20.
10. Narici M, Hoppeler H, Kayser B, Landoni L, Claassen H, Gavardi C, et al. (1996). Human quadriceps cross-sectional area, torque and neural activation during 6 months strength training. Acta Physiol Scand. 1996;157(2):175–86. pmid:8800357
11. Häkkinen K, Pakarinen A, Kraemer WJ, Häkkinen A, Valkeinen H, Alen M. (2001). Selective muscle hypertrophy, changes in EMG and force, and serum hormones during strength training in older women. J Appl Physiol. 2001;91(2):569–80. pmid:11457767
12. Wells AJ, Fukuda DH, Hoffman JR, Gonzalez AM, Jajtner AR, Townsend JR, et al. (2014). Vastus Lateralis Exhibits Non-Homogenous Adaptation to Resistance Training. Muscle Nerve.
13. Butterfield TA, Herzog W. (2006). The magnitude of muscle strain does not influence serial sarcomere number adaptations following eccentric exercise. Pflügers Archiv. 2006;451(5):688–700. pmid:16133258
14. Seynnes OR, de Boer M, Narici MV. (2007). Early skeletal muscle hypertrophy and architectural changes in response to high-intensity resistance training. J Appl Physiol. 2007;102(1):368–73. pmid:17053104
15. Toigo M, Boutellier U. (2006). New fundamental resistance exercise determinants of molecular and cellular muscle adaptations. Eur J Appl Physiol. 2006;97(6):643–63. pmid:16845551
16. Anderson T, Kearney JT. (1982). Effects of three resistance training programs on muscular strength and absolute and relative endurance. Res Q Exerc Sport. 1982;53(1):1–7. pmid:7079558
17. Gentil P, Soares SRS, Pereira MC, d Cunha RR, Martorelli SS, Martorelli AS, et al. (2013). Effect of adding single-joint exercises to a multi-joint exercise resistance-training program on strength and hypertrophy in untrained subjects. Applied Physiology, Nutrition, and Metabolism. 2013;38(3):341–4. pmid:23537028
18. Goto K, Nagasawa M, Yanagisawa O, Kizuka T, Ishii N, Takamatsu K. (2004). Muscular adaptations to combinations of high-and low-intensity resistance exercises. J Strength Cond Res. 2004;18(4):730–7. pmid:15574075
19. Mangine GT, Hoffman JR, Gonzalez AM, Townsend JR, Wells AJ, Jajtner AR, et al. (2015). The effect of training volume and intensity on improvements in muscular strength and size in resistance-trained men. Physiological Reports. 2015;3(8):e12472. pmid:26272733
20. Schoenfeld BJ, Ratamess NA, Peterson MD, Contreras B, Sonmez G, Alvar BA. (2014). Effects of different volume-equated resistance training loading strategies on muscular adaptations in well-trained men. J Strength Cond Res. 2014;28(10):2909–18. pmid:24714538
21. Schoenfeld BJ, Pope ZK, Benik FM, Hester GM, Sellers J, Nooner JL, et al. (2016). Longer Interset Rest Periods Enhance Muscle Strength and Hypertrophy in Resistance-Trained Men. J Strength Cond Res. 2016;30(7):1805–12. pmid:26605807
22. Ema R, Wakahara T, Miyamoto N, Kanehisa H, Kawakami Y. (2013). Inhomogeneous architectural changes of the quadriceps femoris induced by resistance training. Eur J Appl Physiol. 2013;113(11):2691–703. pmid:23949789
23. French D. (2015). Adaptations to anaerobic training programs. In: Haff GG, T T-M, editors. Essentials of Strength Training and Conditioning. 4th ed. Champaign, IL: Human kinetics
24. Berg H, Tedner B, Tesch P. (1993). Changes in lower limb muscle cross-sectional area and tissue fluid volume after transition from standing to supine. Acta Physiologica Scandinavica. 1993;148(4):379–85. pmid:8213193
25. Bemben M. (2002). Use of diagnostic ultrasound for assessing muscle size. Journal of Strength & Conditioning Research. 2002;16(1):103–8.
26. Cadore EL, Izquierdo M, Conceição M, Radaelli R, Pinto RS, Baroni BM, et al. (2012). Echo intensity is associated with skeletal muscle power and cardiovascular performance in elderly men. Experimental Gerontology. 2012;47(6):473–8. pmid:22525196
27. Kumagai K, Abe T, Brechue WF, Ryushi T, Takano S, Mizuno M. (2000). Sprint performance is related to muscle fascicle length in male 100-m sprinters. J Appl Physiol. 2000;88(3):811–6. pmid:10710372
28. Abe T, Brechue WF, Fujita S, Brown JB. (1998). Gender differences in FFM accumulation and architectural characteristics of muscle. Med Sci Sports Exerc. 1998;30(7):1066–70. pmid:9662674
29. Alegre LM, Jiménez F, Gonzalo-Orden JM, Martín-Acero R, Aguado X. (2006). Effects of dynamic resistance training on fascicle lengthand isometric strength. J Sports Sci. 2006;24(05):501–8.
30. Brzycki M. (1993). Strength testing—predicting a one-rep max from reps-to-fatigue. Journal of Physical Education, Recreation & Dance. 1993;64(1):88–90.
31. Cohen J. (1988). Statistical Power Analysis for the Behavioral Sciences 1988. 284–8 p.
32. Weir JP. (2005). Quantifying test-retest reliability using the intraclass correlation coefficient and the SEM. J Strength Cond Res. 2005;19(1):231–40. pmid:15705040
33. Stensdotter A-K, Hodges P, Mellor R, Sundelin G, Häger-Ross C. (2003). Quadriceps activation in closed and in open kinetic chain exercise. Med Sci Sports Exerc. 2003;35(12):2043–7. pmid:14652500
34. Ema R, Sakaguchi M, Akagi R, Kawakami Y. (2016). Unique activation of the quadriceps femoris during single-and multi-joint exercises. Eur J Appl Physiol. 2016;116(5):1031–41. pmid:27032805
35. Luera MJ, Stock MS, Chappell AD. (2014). Electromyographic amplitude vs. concentric and eccentric squat force relationships for monoarticular and biarticular thigh muscles. J Strength Cond Res. 2014;28(2):328–38. Epub 2013/07/31. pmid:23897014.
36. Kawakami Y, Abe T, Fukunaga T. (1993). Muscle-fiber pennation angles are greater in hypertrophied than in normal muscles. J Appl Physiol. 1993;74(6):2740–4. pmid:8365975
37. Kawakami Y, Abe T, Kuno S-Y, Fukunaga T. (1995). Training-induced changes in muscle architecture and specific tension. Eur J Appl Physiol Occup Physiol. 1995;72(1–2):37–43. pmid:8789568
38. Ikegawa S, Funato K, Tsunoda N, Kanehisa H, Fukunaga T, Kawakami Y. (2008). Muscle force per cross-sectional area is inversely related with pennation angle in strength trained athletes. J Strength Cond Res. 2008;22(1):128–31. pmid:18296965
39. Carosio S, Berardinelli MG, Aucello M, Musarò A. (2011). Impact of ageing on muscle cell regeneration. Ageing research reviews. 2011;10(1):35–42. pmid:19683075
40. Lynn R, Morgan D. (1994). Decline running produces more sarcomeres in rat vastus intermedius muscle fibers than does incline running. J Appl Physiol. 1994;77(3):1439–44. pmid:7836150
41. Lynn R, Talbot J, Morgan D. (1998). Differences in rat skeletal muscles after incline and decline running. J Appl Physiol. 1998;85(1):98–104. pmid:9655761
42. Watanabe K, Kouzaki M, Moritani T. (2012). Task-dependent spatial distribution of neural activation pattern in human rectus femoris muscle. J Electromyogr Kinesiol. 2012;22(2):251–8. pmid:22153052
43. Watanabe K, Kouzaki M, Moritani T. (2014). Regional neuromuscular regulation within human rectus femoris muscle during gait. J Biomech. 2014;47(14):3502–8. pmid:25246002
Cita Original
Mangine GT, Redd MJ, Gonzalez AM, Townsend JR, Wells AJ, Jajtner AR, Beyer KS, Boone CH, La Monica MB, Stout JR, Fukuda DH, Ratamess NA, Hoffman JR. (2018) Resistance training does not induce uniform adaptations to quadriceps. PLoS One. 2018 Aug 30;13(8): e0198304. doi: 10.1371/journal.pone.0198304.
Cita en Rev Entren Deport
Gerald T. Mangine, Michael J. Redd, Adam M. Gonzalez, Jeremy R. Townsend, Adam J. Wells, Adam R. Jajtner, Kyle S. Beyer, Carleigh H. Boone, Michael B. La Monica, Jeffrey R. Stout, David H. Fukuda, Nicholas A. Ratamess Jay R. Hoffman (2024). El Entrenamiento de la Fuerza No Induce Adaptaciones Uniformes al Cuádriceps. . (1).https://g-se.com/el-entrenamiento-de-la-fuerza-no-induce-adaptaciones-uniformes-al-cuadriceps-2480-sa-n5c241af7268f8