El Entrenamiento de la Fuerza Hasta el Fallo Mejora la Aptitud Cardiovascular en Humanos: Revisión de las Respuestas Fisiológicas Agudas y Adaptaciones Fisiológicas Crónicas
James Steele1, James Fisher1, Doug McGuff2, Stewart Bruce-Low1, Dave Smith31 Sport Science Laboratory/Centre for Health, Exercise & Sport Science/Southampton Solent University, Southampton, United Kingdom
2 Oconee Medical Centre, Seneca, SC, United States of America
3 Manchester Metropolitan University, Manchester, United Kingdom
Resumen
La investigación demuestra que el entrenamiento de la fuerza produce una mejora significativa en la aptitud cardiovascular (VO2max, economía de movimiento). Hasta la fecha ningún artículo de revisión ha considerado los mecanismos fisiológicos subyacentes que podrían apoyar tales mejoras. Este artículo es una revisión de la narrativa comprensiva, sistemática de la literatura sobre el entrenamiento de la fuerza, aptitud cardiovascular y las respuestas agudas y adaptaciones crónicas que este entrenamiento produce. El principal inconveniente con la investigación existente es la falta de claridad y la cuantificación inadecuada de la intensidad del entrenamiento de la fuerza. Por ello, una consideración importante de esta revisión es el efecto de la intensidad. Las respuestas metabólicas y moleculares agudas frente al entrenamiento de la fuerza hasta el fallo muscular momentáneo no son diferentes a las del entrenamiento de resistencia tradicional. La función del miocardio se mantiene e incluso, quizás aumenta, en la respuesta aguda al entrenamiento de la fuerza de alta intensidad y la intensidad de la contracción parece mediar la respuesta vascular aguda al entrenamiento de la fuerza. Los resultados de adaptaciones fisiológicas crónicas demuestran que el entrenamiento de la fuerza hasta el fallo muscular momentáneo produce varias adaptaciones fisiológicas que pueden facilitar las mejoras que se observan en la aptitud cardiovascular. Las adaptaciones pueden incluir un aumento en las enzimas mitocondriales, la proliferación mitocondrial, la conversión fenotípica de fibras musculares de tipo llx hacia tipo lla, y la remodelación vascular (incluida la capilarización). El entrenamiento de la fuerza hasta el fallo muscular momentáneo produce estímulos agudos suficientes para producir adaptaciones fisiológicas crónicas que mejoran la aptitud cardiovascular. Esta revisión sería la primera en presentar esta conclusión y, por consiguiente, puede ayudar a fomentar un cambio de paradigma que se ocupe del nombre erróneo de ejercicio “cardiovascular” establecido por la modalidad.
Palabras Clave: Aeróbico, Metabólico, Molecular, del miocardio
Tabla de contenidos
1. INTRODUCCIÓN
1.1 MEDICIONES DE ENTRENAMIENTO DE LA FUERZA Y DE APTITUD CARDIOVASCULAR
2. MÉTODOS
2.1 BÚSQUEDA BIBLIOGRAFICA
2.2 CONSIDERACIONES METODOLÓGICAS E INTEPRETATIONES DE LA INVESTIGACIÓN
3. PARTE 1: RESPUESTAS AGUDAS AL ENTRENAMIENTO DE LA FUERZA
3.1 RESPUESTAS DEL COSTO DE OXÍGENO
3.2 RESPUESTAS METABÓLICAS
3.3 RESPUESTAS DEL LACTATO SANGUINEO
3.4 RESPUESTAS MOLECULARES
3.5 RESPUESTAS DEL MIOCARDIO
3.6 RESPUESTAS VASCULARES
4. RESUMEN DE PARTE 1: RESPUESTAS AGUDAS AL ENTRENAMIENTO DE LA FUERZA
5. PARTE 2: ADAPTACIONES CRÓNICAS AL ENTRENAMIENTO DE LA FUERZA
5.1 ADAPTACIONES METABÓLICAS Y MOLECULARES
5.2 ADAPTACIONES DEL MIOCARDIO
5.3 ADAPTACIONES VASCULARES
6. RESUMEN DE PARTE 2: ADAPTACIONES CRÓNICAS AL ENTRENAMIENTO DE LA FUERZA
7. CONCLUSIONES
7.1 FUTURAS ÁREAS DE INVESTIGACIÓN
7.2 APLICACIONES PRÁCTICAS
7.3 CONCLUSIÓN
REFERENCIAS
1. INTRODUCCIÓN
1.1 Mediciones del entrenamiento de la fuerza y de la aptitud cardiovascular.
Artículos de revisión anteriores han examinado los efectos de los pesos libres, máquinas de resistencia variable, máquinas de resistencia hidráulica, máquinas de resistencia neumática sobre la aptitud cardiovascular (80, 144). Para comprender el papel del entrenamiento de la fuerza (RT) en la aptitud cardiovascular (CV), es importante identificar las variables involucradas. Midgley et al. (101) revelaron que las variables de aptitud CV son el consumo de oxígeno máximo (VO2max), economía de movimiento y umbral del lactato (Tlac). La opinión que el RT debe ser complementado con algún tipo de entrenamiento aeróbico o entrenamiento de la resistencia (como correr o ciclismo) para mejorar la aptitud CV está ampliamente aceptada, lo que plantea una dicotomía entre las dos modalidades de entrenamiento. Las revisiones que han analizado los efectos del RT en las variables CV han concluido que aunque el RT puede mejorar tales variables, no es tan eficaz como el entrenamiento aeróbico tradicional o el entrenamiento concurrente (80, 144). Sin embargo, los problemas metodológicos provocan un cierto grado de dificultad en la interpretación. El resultado es que tales declaraciones conclusivas son cuestionables. El problema metodológico mas observado es la inadecuada definición y control de la intensidad del entrenamiento de la fuerza (RT). Recientemente discutimos (48) las definiciones de intensidad junto con los inconvenientes que tiene el uso de porcentaje de 1 repetición máxima (1RM), el cual se refiere a la carga para representar falsamente la intensidad. La intensidad es realmente representativa del esfuerzo, no la carga y por lo tanto sólo es posible una medida exacta durante RT; la de 100%, es decir, cuando el participante alcanza el esfuerzo máximo o el fallo muscular momentáneo (utilizada en esta revisión de manera indistinta con el término “fracaso”). La consideración de este importante factor conduce a una conclusión completamente diferente a la previamente publicada (80, 144).
En los estudios en los que se realizó un control adecuado de la intensidad (definida de esta manera), haciendo que los participantes realizaran RT hasta el fallo muscular momentáneo, los resultados revelaron mejoras tanto en el VO2max estimado (112, 130) como en el VO2max medido en adultos jóvenes (69, 99) y en adultos mayores (67). El trabajo de investigación donde se comparó el RT realizado hasta el fallo, tal como lo recomendamos previamente (48), con el entrenamiento aeróbico, sugirió que las dos modalidades no difieren en el grado de adaptaciones de V02maxque producen (67, 99). Sin embargo, cuando el RT se realizó hasta el fallo, un estudio informó que el RT no mejoró significativamente el VO2max (55). Lamentablemente, los autores no informaron las comparaciones dentro de los grupos y sólo informaron las comparaciones entre los grupos en los datos post entrenamiento entre el grupo control y el grupo que realizó el entrenamiento. Esto significa que no explicaron la diferencia en la aptitud física inicial entre los grupos (que efectivamente era diferente, aunque también se desconoce si era significativa) al comienzo del estudio. Esto podría haber influido en el grado de mejora producida por el entrenamiento. De hecho, tanto el grupo RT como el grupo que realizó ejercicio aeróbico en cinta rodante aumentaron el VO2max. Pero, debido a la falta de comparaciones dentro del grupo, no es posible tener la certeza de que los cambios fueron significativos en el grupo RT. Las revisiones han destacado la falta de evidencia que sustente el uso de RT para mejorar el VO2 en los atletas entrenados (80, 144). Sin embargo, la mayoría de las investigaciones sugieren que es improbable que en una población así se produzcan mejoras significativas en esta variable, independientemente de la modalidad de entrenamiento, debido a su nivel de capacidad de entrenamiento (76).
Una adaptación potencial adicional es la de la economía de carrera (REF), la cual considera el costo de oxígeno en una dada intensidad de ejercicio absoluta. Por ejemplo, si dos individuos realizaran ejercicio con la misma carga de trabajo absoluta, la persona con el menor VO2 sería considerada la mas económica. La mayoría de las investigaciones sobre el efecto de RT en REF indica que los investigadores no han controlado las variables de RT, entre las que se incluye la intensidad (114, 129). También, el uso de programas periodizados hace que sea imposible determinar cuales son las variables que están provocando los efectos observados (45). Sin embargo, estudios en los que el RT se ha realizado hasta el fallo en poblaciones atléticas han reportado una mejora significativa en la economía en comparación con los grupos controles que continuaron realizando el programa de entrenamiento aeróbico habitual (75, 102, 141). Un estudio extenso que analizó el efecto del RT hasta el fallo en la economía en adultos mayores desentrenados observó una mejora significativa en 2 de las 3 tareas funcionales con una disminución significativa en la tasa de intercambio respiratorio. Este hallazgo indica una mayor utilización del metabolismo oxidativo (62).
Otros marcadores de rendimiento CV son el umbral del lactato (Tlac) y la concentración de lactato sub máxima. Se ha demostrado que ambos marcadores mejoran en los sujetos desentrenados como resultado del RT (98). Pero, en los atletas entrenados, Bishop y colegas (19) no observaron ningún cambio significativo en Tlac o en V02. Sin embargo, al igual que el V02, es improbable que el Tlac mejore significativamente en los atletas entrenados (76). Se necesitan mas investigaciones dentro de esta área en las que se contemple un adecuado control de las variables de RT, particularmente la intensidad, y donde se consideren diferentes poblaciones (es decir, sujetos desentrenados y sedentarios).
Los estudios han analizado tanto la metodología de entrenamiento de la fuerza tradicional la cual contempla períodos de descanso que van de 1 a 3 min entre las series y ejercicios (62, 67, 112, 130, 141) y entrenamientos basados en circuitos en los cuales los participantes se mueven rápidamente y hay un descanso mínimo entre los ejercicios (2, 99, 116) y han observado mejoras en la aptitud CV. Además, estos estudios demostraron una variación considerable en el control de otras variables como la carga, volumen y frecuencia. Así, la consistencia entre todos los estudios es que en todos ellos los sujetos realizaron RT hasta el fallo lo que provocó una mejora significativa en las variables de aptitud CV. Parecería que la variable más importante para producir una mejora en la aptitud CV vía RT es la intensidad. O'Hara et al. (111) aportaron una revisión interesante sobre este punto a través del uso de RT para piernas. Los autores concluyeron que el RT de alta intensidad (es decir, hasta el fallo) provocó una marcada mejora en casi todas variables asociadas con la capacidad aeróbica.
En resumen, a pesar de que la investigaciones sobre el efecto de RT sobre el umbral del lactato (Tlac) no han sido concluyentes, parecerían apoyar la recomendación que el RT realizado hasta el fallo (ya sea de tipo circuito o tradicional, e independientemente de otras variables de RT) es suficiente para inducir una mejora significativa en la aptitud CV. Los aumentos en el consumo de oxígeno máximo y en la economía de carrera (RE) pueden ser comparables a los del entrenamiento de resistencia tradicional. Adicionalmente, y corroborando los resultados que el RT realizado hasta el fallo puede producir mejoras en los parámetros de aptitud CV, los estudios también observaron una mejora significativa en el tiempo de resistencia aeróbica hasta la fatiga (2, 69, 116, 141) y en la velocidad en el VO2 (102).
Otros modos de ejercicio realizados en un nivel alto de intensidad también han producido una mejora en la aptitud CV. Por ejemplo, se ha demostrado que entrenamiento intervalado de alta intensidad de ciclismo de ~1,5 hrs/sem produce adaptaciones CV y un aumento en la resistencia similares en a los de un entrenamiento de resistencia tradicional de ~5,5 hrs/sem (27, 53, 66, 88). Dado que la duración sería un factor menos significativo contra la intensidad para producir mejoras en la aptitud a través de esta modalidad, es importante considerar la relevancia de la modalidad y los méritos relativos del RT (notablemente más corto en duración y generalmente superior en intensidad). La dicotomía general que se ha planteado entre el RT y el entrenamiento aeróbico tradicional o entrenamiento de resistencia no tendría fundamento. De hecho, es razonable concluir que la modalidad tendría poca importancia a la hora de producir una mejora en la aptitud CV porque la evidencia indica que esa mejora es posible a través del RT siempre y cuando la intensidad sea alta.
El propósito de esta revisión es presentar los resultados de una búsqueda bibliográfica que sugiere que la dicotomía entre RT y el entrenamiento aeróbico tradicional o entrenamiento de resistencia no es tan clara como se piensa. Esta revisión tiene implicaciones prácticas importantes, particularmente para quienes desean realizar entrenamiento para mejorar la aptitud CV. La revisión comprenderá dos secciones principales. Primero, se examinarán las respuestas fisiológicas agudas al RT que podrían estimular las adaptaciones crónicas para aumentar la aptitud CV. Segundo, esta revisión presentará las investigaciones que han analizado los efectos del RT sobre las adaptaciones fisiológicas crónicas, y se considerarán los mecanismos del RT que podrían ser los responsables de la estimulación de tales adaptaciones.
2. MÉTODOS
2.1 Búsqueda de literatura
Antes de escribir esta revisión se realizó una búsqueda de literatura entre marzo de 2010 y enero de 2011 en bases de datos como MEDLINE/Pubmed, SportDiscus y el buscador Google académico, utilizando diferentes palabras claves asociadas con el RT, la aptitud CV y variables/procesos fisiológicos (por ejemplo, “RT y CV”, “RT y resistencia”, “RT y aeróbico”, “RT y VO2”, “RT y economía”, “RT y umbral del lactato”, “RT y metabólico”, “RT y molecular”, “RT y del miocardio”, “RT y vascular” así como otros sinónimos asociados, palabras y combinaciones similares). El entrenamiento de la fuerza engloba apreciablemente una amplia gama de tipos de fuerza, y se reconoce que de hecho todo el movimiento ocurre contra una resistencia tanto interna como externa. Sin embargo, el estilo de rendimiento físico independiente de los tipos de resistencia sería de importancia clave(48) y por lo tanto, dentro de esta revisión, el RT como modalidad de ejercicio fue considerado como aquel ejercicio que utiliza cualquiera de los siguientes tipos de resistencia típicos: pesos libres (incluidos aquellos ejercicios con peso corporal realizado de la misma manera característica), máquinas de resistencia variable, máquinas de resistencia hidráulica y máquinas de resistencia neumática. Los estudios que contemplaban entrenamiento de “alta resistencia”, pero fueron realizados en una bicicleta ergométrica no fueron considerados como RT. En cambio, fueron considerados como modalidades de entrenamiento de resistencia tradicional. Inicialmente, los criterios de inclusión y exclusión se centraron en metodologías que comprendieran estudios controlados y aleatorizados que analizaran las respuestas agudas a un estímulo de RT, así como también estudios controlados de intervención de entrenamiento que analizaran los fenómenos fisiológicos en respuesta a RT. Sin embargo, como el número de artículos inicialmente excluido creció, fue necesario evaluar nuevamente el proceso. Se evidenció que muchos trabajos contenían numerosas inconsistencias metodológicas que dificultaban una interpretación concisa, las cuales están documentadas detalladamente en el artículo presente. Como resultado, se decidió tomar el enfoque de una revisión narrativa sistemática que utilizó una búsqueda específica y métodos de inclusión, pero no especificamos métodos para la apreciación crítica de los estudios incluidos. Esto permitió una interpretación holística de las variadas y de las numerosas investigaciones identificadas. También permitió los beneficios de utilizar ambos tipos de revisiones (36). Nosotros limitamos nuestro criterio de inclusión a aquellas investigaciones que involucraran poblaciones saludables, desentrenadas y atléticas que hubieran analizado: (a) variables agudas que se consideraban involucradas en la mejora de la aptitud CV en respuesta al RT; (b) medidas de aptitud CV en respuesta a una intervención de RT (por ejemplo, VO2max, tiempo de resistencia hasta la fatiga, umbral del lactato y economía de movimiento); y (c) la adaptación en variables fisiológicas que se consideraba que podían contribuir con una mejora en la aptitud CV (y destacadas en las consideraciones metodológicas) en respuesta al RT.
2.2 Consideraciones metodológicas e interpretaciones de las investigaciones
Tal como resaltamos previamente, la definición y el control de la intensidad fueron considerados importantes para la interpretación de los resultados. Por tal motivo registramos aquellos estudios que realizaron un control de estos factores mientras se realizaba el RT hasta el fallo. Respecto a los estudios que no definieron o controlaron la intensidad pero sus participantes realizaron RT de alta intensidad con resultados potencialmente importantes, presentaremos una interpretación razonable de la intensidad del RT. Aunque esto se basó en el conocimiento de los rangos de una repetición máxima típica con cargas particulares relativas (70, 133), reconocemos la limitación de cuantificar los resultados sobre la base en nuestra propia definición. Se sugiere que en las futuras investigaciones la definición y la intensidad se establezcan apropiadamente para que se pueda obtener una interpretación más clara de los resultados.
Al determinar las respuestas agudas y las adaptaciones crónicas que debíamos analizar, quedaba claro que existía mucha incongruencia con respecto a los factores más importantes involucrados con la determinación de las medidas de rendimiento de resistencia y aptitud CV (15, 16, 106, 107, 108, 123). Dado que no era el propósito de esta revisión discutir la validez de una o de otra hipótesis, se analizaron áreas seleccionadas de adaptación que se consideraba que tenían el mérito de mejorar la resistencia CV. Estas fueron: (a) CV (es decir, morfología del miocardio y morfología vascular); (b) metabólica (es decir, enzimas oxidativas); y (c) molecular (es decir, composición de la fibra muscular y densidad mitocondrial). También se analizaron las respuestas agudas que podían ser consideradas importantes para la estimulación de las adaptaciones que permitieran un aumento en la aptitud CV (es decir, el costo de oxígeno, tensión arterial, frecuencia cardíaca, volumen de sangre bombeado y las respuestas vasculares), metabólicas (es decir, actividad metabólica anaeróbica, actividad metabólica aeróbica y lactato sanguíneo), y molecular (i.e. las vías de señalización molecular). Se presentaron adaptaciones crónicas como adaptaciones CV vinculadas al sistema de transporte de oxígeno (es decir, adaptaciones del miocardio y adaptaciones vasculares), y adaptaciones metabólicas y moleculares a nivel muscular local (entre las que se incluían adaptaciones enzimáticas, el fenotipo de las fibras musculares y adaptaciones mitocondriales). Siguiendo lo establecido en estas distinciones también se presentaron los correspondientes estímulos agudos.
Para obtener las conclusiones, se utilizó la metodología de “peso de la evidencia disponible” para proporcionar una interpretación holística de la investigación y plantear hipótesis sobre los mecanismos fisiológicamente creíbles para explicar el efecto de RT hasta el fallo sobre las variables de aptitud CV.
3. PARTE 1: RESPUESTAS AGUDAS AL RT
3.1 Respuestas del Costo de Oxígeno
Para estimular una mejora en la resistencia CV, los sujetos desentrenados y los deben trabajar a ~50% y ~70 a 80% del VO2max respectivamente (9). Por lo tanto, la pregunta que surge es: ¿"Este pensamiento también se puede aplicar al RT? ". Podría presumirse que dado que el RT hasta el fallo provoca un aumento significativo en las medidas de aptitud CV, también debe provocar una respuesta significativa en el VO2. Por lo tanto es interesante examinar si éste puede ser un estímulo agudo durante RT que pueda estimular una mayor aptitud CV. Lamentablemente, al examinar el VO2 del RT, muchos estudios o no controlaron la intensidad o no la definieron suficientemente (18, 56, 124). Como resultado, es cuestionable si los resultados representan la respuesta del VO2 con precisión durante el RT hasta el fallo. Otros estudios han realizado RT hasta el fallo, pero los datos fueron registrados como promedio a lo largo de una sesión e incluyendo los períodos de descanso por lo que sólo una pequeña fracción de tiempo representaba lo realmente destinado al RT (~6.21 a 8 min de ejercicio de los 24 a 30 min medidos (20, 117, 118). De hecho, Phillips y Ziuraitis (117, 118) informaron una intensidad moderada de RT basada en equivalente metabólicos (METS) de 3 a 6 para las sesiones de RT, a pesar de que los sujetos realizaron ejercicio hasta el fallo (es decir, intensidad máxima). El VO2 y otras variables CV podrían haber sido significativamente infra valoradas cuando se efectuó el promedio. Se sabe que el índice de esfuerzo percibido (RPE) es significativamente menor cuando los datos se promedian incluyendo los períodos de descanso (143) de la misma manera que los estudios mencionado de VO2. Esto sugiere que es probable que la respuesta del VO2 observada en estos estudios no haya reflejado con precisión la intensidad del RT.
También se ha demostrado que el VO2 de un ejercicio particular depende de la masa muscular activa (9, 142) algo que debe ser considerado cuando se mide el VO2 en los estudios de RT. La mayoría de los estudios han informado que el VO2 promedio durante RT es inferior al 50% de la capacidad corporal total máxima (17, 20, 28, 35, 43, 74, 117, 118). Así, en función de las recomendaciones establecidas para alcanzar niveles particulares de VO2 con el fin de mejorar la aptitud CV, el VO2 promedio durante el RT es el dato que permitió a Jung (80) concluir que el VO2 del RT es "... apenas un estímulo sustancial para mejorar la capacidad aeróbica en todos, excepto en las personas sedentarias." Sin embargo, la investigación realizada por Stromme et al. (142) y los datos recolectados por Astrand et al. (9), sugieren que el VO2 durante RT debe ser relativamente menor en comparación al de los ejercicios que involucran mayor masa muscular como los ejercicios en cinta rodante o el esquí de fondo (sobre todo si se mide de manera aislada para un ejercicio específico). Tal vez, al considerar el VO2 durante el RT, debería analizarse en relación al VO2 máximo alcanzado durante el RT en oposición al VO2 máximo del cuerpo entero obtenido con ejercicios que involucran una mayor masa muscular.
A pesar de las dificultades para determinar el VO2 durante el entrenamiento de la fuerza (RT) de alta intensidad, es razonable esperar que cuando se incremente la intensidad del RT se produzca un aumento concomitante en VO2. De hecho, Gotshalk et al. (56) demostraron que durante una sesión en forma de circuito de ejercicios de peso en la cual se mantuvieron constantes la carga y las repeticiones a lo largo de los circuitos completados, el VO2 aumentó en cada circuito, probablemente a medida que los sujetos experimentaban fatiga. A medida que aumentaba el grado de esfuerzo, aumentaba la intensidad del ejercicio realizado. También se ha sugerido que las repeticiones de mayor duración pueden tener una intensidad relativamente más alta que las de menor duración (25). Por consiguiente, nuevamente, es razonable esperar que la mayor intensidad producida por las repeticiones más largas provoque una mayor respuesta de VO2. Barreto et al. (14) no informaron ninguna diferencia en los valores de VO2 en ejercicios de circuitos de repetición de mayor duración (2 seg fase concéntrica y 2 seg fase excéntrica) en comparación con los de menor duración (1 seg de fase concéntrica y 1 seg de fase excéntrica). Sin embargo, los autores también comentaron que la diferencia en las duraciones de las repeticiones podría no haber sido suficiente para demostrar una diferencia significativa en VO2. Hunter et al. (73) efectuaron una comparación entre entrenamiento súper lento (SS) (10 seg fase concéntrica, 5 seg fase excéntrica) y entrenamiento tradicional de RT (TT). No se aplicó ninguna restricción en la duración de TT (duración promedio de la fase concéntrica fue 0,9 seg y el promedio de la excéntrica fue 0,8 seg). Inicialmente, sus resultados sugirieron que el TT provocó un aumento mayor en VO2 que el SS. Sin embargo, el grupo TT entrenó con una carga significativamente mayor (65% 1RM) que la del grupo SS (25% 1RM) durante un período idéntico de tiempo, lo que sugiere que el grupo TT podría haber entrenado a una intensidad más alta (es decir, más cerca del fallo).
No queda claro a partir de la investigación actual si la verdadera respuesta de VO2 fue a partir del RT realizado hasta el fallo. Esto se debe principalmente a problemas metodológicos como promediar el VO2 en las sesiones de RT y, luego, compararlo con el VO2 del cuerpo entero determinado con ejercicios para mayor masa muscular. El aumento en la intensidad durante RT puede aumentar la respuesta del VO2, que hipotéticamente podría incluir un aumento en la duración de la repetición. Sin embargo, quizás debido a las diferencias insuficientes en la duración de los protocolos y al control inadecuado de la intensidad, esto no se demuestra en las investigaciones publicadas. Los estudios futuros deben usar diseños de investigación que permitan evaluar la hipótesis que las repeticiones de mayor duración producen una mayor respuesta de VO2 debido a una intensidad más alta (es decir, manteniendo la carga y la duración de la serie constantes y sólo variando la duración de la repetición entre las condiciones). Entonces, además, ¿por qué no utilizar un diseño de investigación que controle la intensidad del RT y registre el VO2 agudo durante RT y no obtener un promedio en las sesiones donde se incluye el descanso? Esto ayudaría a aclarar si la respuesta del VO2 durante RT es un estímulo importante para las adaptaciones de aptitud CV. De hecho, un concepto erróneo común es que se requiere un elevado porcentaje de VO2max de cuerpo entero para aumentar la capacidad aeróbica. De hecho, una revisión de la literatura demuestra que no hay ninguna evidencia sustancial que sugiera que sea necesario un porcentaje definido de VO2max de cuerpo entero durante un típico programa de entrenamiento de la resistencia para aumentarla (101). El estado de entrenamiento de los músculos activos también influye en la determinación del VO2 (68, 142) lo que indica una adaptación a nivel muscular independiente del VO2 del cuerpo entero obtenido durante el ejercicio de resistencia. Esto sugiere que considerar el VO2 durante el entrenamiento medido en relación al VO2max obtenido a través de ejercicios que involucren una gran masa muscular, como indicador de su eficacia para mejorar la aptitud CV puede tener poco valor.
3.2 Respuestas metabólicas
Se sabe que el entrenamiento aeróbico es diferente al entrenamiento anaeróbico. El primero es considerado sinónimo de entrenamiento de resistencia CV o tradicional mientras que el segundo es sinónimo de entrenamiento con sobrecarga o entrenamiento de fuerza de alta intensidad (154). Sin embargo existe una probabilidad muy real, que la distinción entre las dos condiciones sea una simplificación excesiva dónde tal dicotomía no existe de hecho. Las definiciones implican que el entrenamiento anaeróbico es un estímulo insuficiente para mejorar el metabolismo aeróbico. De hecho, muchos investigadores concluyeron que el RT no es el estímulo apropiado para mejorar el metabolismo aeróbico (17, 20, 28, 35, 43, 74, 80, 117, 118, 143). Esta conclusión parece provenir de los conceptos enseñados tradicionalmente sobre cada tipo de entrenamiento. ¿Pero, que pasaría si esta creencia es un concepto erróneo infundado en que la energía que producen las vías metabólicas involucradas en la contracción muscular pueden ser diferentes en función de la modalidad de ejercicio (37) y no en función de la intensidad? Metabólicamente, el RT debería estimular el sistema aeróbico.
Aunque el concepto de “aeróbico” fue popularizado por Cooper (37) para referirse al uso aislado de las vías aeróbicas para producir aumentos en la resistencia CV, la potencia de las células (mitocondrias) se alimenta con piruvato producido por una serie de pasos químicos que no requieren oxígeno (glucólisis). Esto significa que el metabolismo anaeróbico y el metabolismo aeróbico están vinculados metabólicamente (127). Además, es muy probable que la glucólisis aeróbica (o sea el ingreso de piruvato al Ciclo de Krebs cuando hay un nivel de oxígeno adecuado que sostenga su vinculación con el sistema de transporte de electrones) sea estimulada por el ejercicio aeróbico de alta intensidad independientemente de la modalidad. En concordancia con esto, numerosos estudios han demostrado una mejora en la capacidad aeróbica luego de entrenamiento intervalado de alta intensidad (27, 53, 66, 88). Es probable que se produzcan adaptaciones similares a través del RT hasta el fallo y por ello se sugiere que la modalidad de ejercicio es irrelevante para el estímulo de las vías metabólicas aeróbicas.
El efecto de la intensidad es especialmente importante en la discusión de si el RT produce adaptaciones aeróbicas. Una intensidad de ejercicio creciente provoca un aumento en el reclutamiento de unidades motoras hasta un máximo regulado, el cual es independiente de la modalidad del ejercicio (30, 83, 110). También, probablemente, la piruvato deshidrogenasa tendrá una actividad máxima cuando la intensidad del ejercicio sea máxima. Esto es importante porque es la enzima limitante de la velocidad, responsable de la entrada de piruvato en la mitocondria (135). Mayer y colegas (92) demostraron que durante el ejercicio máximo, la infusión de dicloroacetato, que es un estimulador metabólico de la piruvato deshidrogenasa producía un aumento en el VO2. En otras investigaciones se aplicó infusión intra-arterial de adenosina para inducir una respuesta vasodilatadora en los cuadriceps mientras se realizaban ejercicios máximos de extensión de rodilla con una sola pierna (13) y un test máximo en bicicleta ergométrica (29) para inducir una mayor entrega de O2, sin embargo no se observaron aumentos en el VO2 muscular ni en el VO2 del cuerpo entero. Esto apoya el concepto que el trabajo muscular de máxima intensidad, independientemente de la modalidad, puede producir tasas máximas de metabolismo aeróbico en la musculatura activa local tal como se determina por la tasa de entrada del piruvato a la mitocondria por la piruvato deshidrogenasa. Esto se demuestra por el hallazgo que el suministro de oxígeno es aparentemente adecuado para alcanzar la intensidad. Por consiguiente, aplicado a RT (13), esto sugiere que el entrenamiento hasta el fallo también provoca la mayor actividad aguda tanto de la glucólisis anaeróbica como de la glucólisis aeróbica en la musculatura activa local, y tal entrenamiento probablemente es un estímulo suficiente para la energía aeróbica (43) y la adaptación.
3.3 Respuestas del lactato sanguíneo
La discusión sobre las respuestas metabólicas al RT incluye el tema del lactato sanguíneo (Blac) debido a que en concentraciones altas, tendría efectos potencialmente perjudiciales sobre la contracción y resistencia muscular a través de la acidosis metabólica subsiguiente (77, 154). Se ha sugerido que una mayor actividad de las enzimas oxidativas asociada el entrenamiento de resistencia provoca una mayor tasa de entrada de piruvato a la mitocondria y una menor acción de la lactato deshidrogenasa (77). Sin embargo, tal como se explicó previamente, la enzima piruvato deshidrogenasa que es la responsable del transporte de piruvato hacia la mitocondria, es la enzima limitante de la velocidad (135). Por lo tanto cualquier adaptación en el Blac producida por el RT probablemente se deba a la estimulación y a la mayor expresión de los transportadores de monocarboxilato que facilitan una mejor remoción del lactato, lo que es similar al efecto que produce el entrenamiento de resistencia (42).
Se ha demostrado que las respuestas del lactato sanguíneo (Blac) aumentan con la intensidad progresivamente mayor de la contracción muscular (72, 134). Esto es esperable debido a la naturaleza limitante de la velocidad de la enzima piruvato deshidrogenasa (135). La glucólisis anaeróbica alcanza los requerimientos energéticos y como consecuencia produce piruvato a una velocidad superior a la del transporte de piruvato hacia la mitocondria y en consecuencia el piruvato y los iones hidrógeno incrementan su concentración en el citoplasma. La enzima lactato deshidrogenasa cataliza la reacción entre las concentraciones crecientes de estos productos de la glucólisis anaeróbica para producir lactato. Como el RT hasta el fallo es un estímulo máximo tanto para el metabolismo aeróbico como para el metabolismo anaeróbico, podría esperarse que se incremente la producción de lactato sanguíneo durante el ejercicio y, como consecuencia, se estimulen los mecanismos involucrados con su remoción y metabolismo.
Tal como se discutió para otras variables, la intensidad debe ser definida con precisión y debe ser controlada en los trabajos de investigación. Los estudios que han utilizado RT hasta el fallo observaron aumentos significativos en el lactato sanguíneo (Blac) agudo (~6,5to 17 mmol L-1) (31, 39, 52, 74, 148). Sin embargo, está claro que los protocolos utilizados en estos estudios varían considerablemente (es decir, carga, series, repeticiones y rango) lo que dificulta el análisis de cualquier variable de RT particular que este ejerciendo un efecto en la acumulación de Blac a excepción de la intensidad. Adicionalmente, el estudio de Dentón y Cronin (39) requiere una interpretación cuidadosa porque compararon 3 esquemas diferentes de descanso y carga, y solo en uno de ellos los participantes realizaron el ejercicio hasta el fallo. De hecho, éste fue el único grupo que presentó un aumento significativo en Blac.
Las repeticiones de mayor duración pueden tener mayor intensidad que las de menor duración (25). La dificultad para interpretar los resultados de Hunter et al. (73) fue descripta en la sección sobre VO2, algo que también podría aplicarse a los hallazgos sobre el Blac con resultados que favorecen a grupo que entrenó a una intensidad mayor (grupo TT) porque presentó una mayor acumulación de Blac. Gentil y colegas (52) también analizaron el entrenamiento súper lento (SS) y concluyeron que aporta poco como estímulo metabólico. Los autores realizaron una comparación entre 4 grupos que realizaron ejercicios de extensión de piernas con 10 RM. El primer grupo realizó las repeticiones con una duración SS, el segundo grupo realizó las repeticiones tradicionales, y los dos grupos restantes realizaron diferentes contracciones isométricas con extensión completa. Todos los grupos excepto el grupo SS, realizaron las repeticiones hasta el fallo definidas como "... cuando el sujeto no pudo extender las rodillas completamente durante dos repeticiones consecutivas". El grupo SS realizó una sola repetición estandarizada de 60 seg. con 30 seg para la fase excéntrica y 30 seg para la frase concéntrica, y por lo tanto no podemos saber si fue hasta el fallo y, por consiguiente, de intensidad máxima. Los resultados revelan que no se observó ninguna diferencia entre los grupos que realizaron ejercicio hasta el fallo, y solo se observaron diferencias significativas entre los grupos que realizaron los ejercicios isoméricos y el grupo SS.
El ejercicio que produce una acumulación de lactato sanguíneo igual o superior a la del umbral del lactato (Tlac) produce una mejora significativa en Tlac (38, 47). Como mencionamos previamente (31, 39, 74, 148), el RT produce concentraciones relativamente altas de lactato en sangre (~6,5 a 17 mmol L-1). Parecería que el RT podría producir un aumento en los mecanismos de aclaramiento de lactato. Pero a pesar de la plausibilidad intuitiva de esta hipótesis, se necesitan más investigaciones en forma de intervenciones de entrenamiento controladas para confirmarla.
3.4 Respuestas moleculares
Las vías de señalización molecular controlan muchas de las adaptaciones potencialmente estimuladas. Recientes revisiones (11, 32, 85, 105) refuerzan adicionalmente la dicotomía entre las adaptaciones en la aptitud CV producidas por el entrenamiento RT y por el entrenamiento tradicional. El argumento que las adaptaciones de resistencia son mutuamente exclusivas de las adaptaciones RT es específico de lo que Atherton et al. (10) han denominado el switch AMPK-PKB. La vía de la proteína quinasa activada por monofosfato de adenina (AMPK) desempeña un papel importante en: (a) la inducción de la biogénesis mitocondrial (es decir, proliferación mitocondrial y la regulación de enzimas mitocondriales) (122 ,157); (b) estimulación de la transformación del fenotipo de fibras de contracción lenta (91); y (c) inducción de la formación de propiedades oxidativas en las fibras de tipo llx (5). Así, la vía AMPK se postula como el instigador fundamental de las adaptaciones de resistencia en el músculo esquelético.
En cambio, en los mamíferos el blanco de la vía de la rapamicina (mTOR) induce una cascada de eventos que producen un aumento en la síntesis de proteínas musculares (es decir, hipertrofia) (21, 22). Atherton et al. (10) analizaron el efecto de dos programas de estímulos eléctricos diferentes en el músculo de ratas; un programa de alta frecuencia que supuestamente simulaba el RT, y un programa de baja frecuencia que simulaba el entrenamiento resistencia. Los resultados indicaron que el ejercicio de resistencia favorecía la activación de AMPK y el ejercicio de fuerza favorecía la activación de mTOR. Los dos tipos de ejercicio serían mutuamente excluyentes. Es importante destacar que los trabajos de revisión sobre este tema (11, 32, 85, 105) concluyeron que los ejercicios de resistencia y de fuerza son diferentes en su señalización molecular aguda y en sus adaptaciones crónicas. Los dos utilizan el switch AMPK-PKB descripto por Atherton et al. (10). Sin embargo, es muy probable que estas conclusiones sean algo tenues cuando se consideran las limitaciones metodológicas del estudio de Atherton et al. (10), particularmente respecto a los participantes no humanos y a la deficiente simulación de programas de entrenamiento humanos por medio de un estímulo eléctrico.
Otros estudios también han demostrado que el AMPK sería un inhibidor de la activación de mTOR. Hardie y Sakamoto (60) presentaron una revisión comprensiva sobre este tema. Luego, para aclarar también este tema Dreyer et al. (41) en un estudio realizado con personas, demostraron que durante el RT de alta intensidad (10 series de 10 repeticiones a 70% de 1RM pero debieron reducir la carga para algunos participantes para lograr 10 repeticiones, lo que sugiriere que se alcanzó el fallo), la AMPK fue activada significativamente y mTOR fue inhibido. Este hallazgo no coincide con las sugerencias basadas en el estudio de Atherton et al. (10) que el RT no activa AMPK. Dreyer et al. (41) también informaron que 1 a 2 hrs después del RT los reguladores corriente arriba de mTOR, como la fosforilación de la proteína quinasa B (PKB; la activación de PKB desactiva el complejo de esclerosis tuberosa tipo 2, un inhibidor del proceso mTOR corriente abajo) y la fosforilación del mTOR aumentaron significativamente. Este estudio sugiere el potencial del RT para producir adaptaciones asociadas con el entrenamiento de resistencia. El estudio de Koopman et al. (84) apoya estos resultados. Estos investigadores usaron un protocolo similar de RT. Por otra parte, los efectos agudos de activación de AMPK sólo inhiben el mTOR durante el ejercicio (21) mientras que se produce una regulación por incremento transitoria post ejercicio de los reguladores corriente arriba de mTOR y se observa un aumento retardado en la fosforilación de mTOR (21, 41).
Frosig et al. (51) analizaron la actividad de AMPK en respuesta al entrenamiento de extensión de piernas. Aunque lo describieron como un protocolo de entrenamiento de resistencia, el entrenamiento se realizó a 100% de trabajo máximo con el fin de "... asegurar el reclutamiento de la mayoría de las fibras musculares... ". Esto podría ser considerado RT de alta intensidad sobre la base de la definición de intensidad usada en esta revisión. Los autores también informaron un aumento significativo en la actividad de AMPK. La implicación es que el ejercicio realizado a una intensidad lo suficientemente alta produce activación de la vía de AMPK independientemente de la modalidad del ejercicio (41, 51). En concordancia Gibala et al. (54) demostraron que el AMPK también se activa durante el ejercicio intervalado de alta intensidad. Que la AMPK se active independiente de la modalidad siempre que la intensidad sea alta no es sorprendente porque la AMPK es un sensor fundamental de requerimientos energéticos celulares y responde rápidamente a los aumentos en la relación celular AMP:ATP (59, 60). Durante el ejercicio de alta intensidad la relación AMP:ATP aumenta debido a la mayor proporción de uso de ATP (72, 128, 136) y así se activa la AMPK. El reclutamiento de fibras musculares de tipo llx durante RT hasta el fallo (30) produce un mayor agotamiento de ATP debido a la mayor actividad ATPasa de la miosina de las fibras (128). Esto coincide con el hallazgo que la activación de AMPK también es mayor en las fibras de tipo llx después del ejercicio (89). El entrenamiento de la fuerza hasta el fallo desencadenará la activación de AMPK a través de estos procesos, así como la activación retardada subsiguiente de mTOR, lo que presenta un mecanismo molecular a través del cual el RT puede producir una mejora en la aptitud CV, en la fuerza y en la hipertrofia. Por último, activación de las células satélite a través del RT también induce cambios en los genes mitocondriales (147) lo que probablemente también controlaría la adaptación mitocondrial en respuesta al RT.
3.5 Respuestas del miocardio
El sistema de transporte de oxígeno y por lo tanto, el VO2 y sus constituyentes metabólicos han sido discutidos en parte. Sin embargo, tal como señalamos en la sección de métodos, el sistema de transporte de oxígeno también está influenciado por las variables CV centrales. Una discusión sobre las respuestas agudas que podrían estimular la adaptación en las variables CV centrales es importante para esta revisión. Por ejemplo, se piensa que las respuestas de incremento en la presión arterial con el RT podrían estar relacionadas con la adaptación en las dimensiones cardíacas (33). La adaptación podría producir una mejora en el gasto cardíaco máximo y en la entrega de oxígeno.
Se ha demostrado previamente que el entrenamiento de la fuerza está asociado con un aumento en la presión arterial sistólica (SBP) en un rango de 270 a 480 mmHg (8, 94, 96). En general, los estudios demuestran que el RT aumenta significativamente la presión sanguínea (BP) y el efecto más pronunciado se observa en la SBP (8, 81). El efecto sobre la presión arterial diastólica (DBP) es menos pronunciado y, en algunos casos, no cambia o disminuye ligeramente (8, 81). El tipo de contracción muscular también afecta la respuesta de presión arterial. Okamoto et al. (113) observaron que cuando la carga se fijaba en relación a 80% de torque máximo, las contracciones concéntricas producían una respuesta mayor a las observadas durante las contracciones excéntricas en todas las variables CV medidas. Esto no es sorprendente porque las contracciones concéntricas también provocan una mayor respuesta de VO2 (7) debido a la necesidad de activación de más unidades motoras para producir la misma producción de fuerza que la contracción excéntrica (78). De manera similar, Huggett et al. (71) demostraron que las contracciones isométricas provocaban una respuesta hemodinámica mayor que las contracciones excéntricas. Okamoto et al. (113) también demostraron un aumento en las variables hemodinámicas que incrementaban el consumo de oxígeno del miocardio a medida que aumentaba la intensidad del ejercicio concéntrico.
Los cambios en la presión arterial durante el ejercicio deben distinguirse entre las respuestas que ocurren a nivel del miocardio y las respuestas que ocurren a nivel de la vasculatura periférica. De hecho, podrían producirse efectos adaptativos separados centralmente y periféricamente. Esto es porque el aumento en SBP durante RT es secundario a la mayor presión intratorácica asociada con una maniobra del valsalva (90, 94), y la mayor presión intratorácica que se transmite directamente a la vasculatura arterial a medida que aumenta la SBP (65). Así, la presión a la que el miocardio es “expuesto” (es decir, presión transmural del ventrículo izquierdo = presión del ventrículo izquierdo – presión intratorácica) no se eleva por encima de los valores de reposo (26). Esto fue corroborado adicionalmente por Haykowsky et al. (63) quienes informaron que mientras la presión intratorácica aumentó significativamente de 1,7 mmHg a 111,7 mmHg durante un ejercicio de press de piernas a l80% 1RM, no se observó ninguna diferencia significativa en el estrés de la pared del ventrículo izquierdo (LV).
Cuando los cambios hemodinámicos centrales se miden directamente por medio de cateterización en la parte derecha del corazón durante RT, se observa una disminución significativa en la resistencia vascular periférica y mayores índices de trabajo cardíaco e índices de trabajo sistólico del ventrículo izquierdo (100). Estas respuestas son indicativas de una mayor perfusión coronaria. Los ecocardiogramas también demuestran que la función del ventrículo izquierdo se mantiene durante RT en los sujetos saludables (82). A medida que se incrementa la intensidad del RT, también aumenta de manera lineal la velocidad del flujo sanguíneo (3, 46). Las respuestas hemodinámicas centrales al RT (es decir, ningún cambio significativo en la presión cardíaca directa) se deberían en parte a la acción de bombeo del músculo esquelético que refuerza el retorno venoso (140). La falta de una acción de bombeo por parte del músculo esquelético podría explicar la mayor respuesta hemodinámica en las contracciones isométricas (71). Es posible aumentar el flujo de sangre y la perfusión coronarios debido al mayor retorno venoso (44), que produce una mayor actividad del ventrículo izquierdo durante el RT.
Estos resultados parecen indicar que existe un estímulo agudo pequeño que produce adaptación en la dimensión del miocardio. Sin embargo, la frecuencia cardíaca (HR) también aumenta significativamente durante RT (8, 71, 81, 113). Como ocurre con otras variables fisiológicas, parecería que cuando se controla la intensidad, la HR aumenta linealmente. Arent et al. (6) demostraron que cuando se controla el volumen, un aumento en la carga que lleva al sujeto a realizar ejercicio hasta el fallo produce un aumento lineal en la HR y en el índice de esfuerzo percibido (RPE). El mayor estrés en el miocardio por un aumento en la HR podría ser posiblemente uno de los mecanismos mediante los cuales se produce la adaptación. Alternativamente, el volumen de sangre bombeado por el corazón también sería un posible mecanismo para la adaptación. Morganroth et al. (104) sugirieron que la adaptación de la hipertrofia excéntrica (es decir, aumento proporcional en el tamaño de la cámara del ventrículo y espesor de la pared del ventrículo izquierdo) es el producto de un rendimiento cardíaco alto mientras que la hipertrofia concéntrica caracterizada por una mayor relación espesor de la pared:radio sería el resultado del volumen de sangre bombeado, que podría ser mayor en los entrenamientos de alto volumen preferidos por los fisiculturistas en contraste con las sesiones de carga potencialmente más altas de los levantadores de pesas. En concordancia con esto, Haykowsky et al. (64) informaron que la hipertrofia concéntrica esta asociada a levantadores de pesas olímpicos mientras la hipertrofia excéntrica se asocia con los fisiculturistas. Aunque ésta es una especulación basada en una asociación, será discutida en la sección posterior sobre las adaptaciones del miocardio a RT.
3.6 Respuesta vascular
El mayor gasto cardíaco mejora el transporte de oxígeno mientras que el suministro de sangre local al músculo afecta directamente la entrega de oxígeno. Por consiguiente, la estimulación aguda de la vasculatura periférica podría resultar en un mayor suministro de oxígeno local. Brown (24) sugirió que los estímulos agudos primarios que determinan las adaptaciones de remodelación vascular son la tensión de cizallamiento y tensión de la pared de la vasculatura. Como explicamos, a mayor intensidad de RT, la velocidad del flujo sanguíneo aumenta linealmente (3, 46). Tinken et al. (150) observaron que la tasa de cizallamiento aumentaba significativamente en respuesta a ejercicios de RT para puños. Un aumento en el flujo sanguíneo y por consiguiente en la tensión de cizallamiento debida al RT puede aportar un estímulo para la adaptación endotelial en la producción de óxido nítrico (un potente vasodilatador que puede servir para aumentar el flujo de sangre local). Shen y colegas (132) identificaron la tensión de cizallamiento como el estímulo principal para la producción de óxido nítrico por parte del endotelio. Los autores también comentaron que el ejercicio breve puede regular positivamente (regulación por incremento) la expresión génica de la enzima responsable de su producción. Sin embargo, carecemos de datos obtenidos en humanos que puedan demostrar esta hipótesis. Se ha demostrado que el entrenamiento de resistencia de larga duración realizado a baja intensidad no sería eficaz para alterar los niveles plasmáticos de nitrato/nitrito (23). Sin embargo, habría algún efecto del entrenamiento en la respuesta vasodilatadora tal como se observa en la mayor magnitud de bloqueo en la vasoconstricción inducida por frío en ciclistas competitivos en comparación con los controles sedentarios cuando ambos grupos realizan ejercicios de extensión de piernas o de puño (156). Además, Wray et al. (156) observaron que la conductancia vascular en respuesta al ejercicio máximo hasta la fatiga aumentaba significativamente tanto en los sujetos sedentarios como en los sujetos entrenados en resistencia, pero la respuesta fue mayor en los sujetos que tenían entrenamiento en resistencia. Es importante que se realicen más investigaciones para identificar si esta respuesta también se produce en los seres humanos como resultado de RT hasta el fallo. DeVan y colegas (40) observaron una disminución en la elasticidad arterial que aumentaría el flujo de sangre luego del RT hasta el fallo. Sin embargo, la disminución duró menos de 60 min después de la realización del ejercicio. En función de este punto y del aumento dependiente de la intensidad de la contracción del flujo de sangre (3, 46), las futuras investigaciones deberían analizar si las adaptaciones vasculares debidas al RT hasta el fallo están efectivamente reguladas por este mecanismo.
4. RESUMEN DE LA PARTE 1: RESPUESTAS AGUDAS AL RT
Como hemos observado, las respuestas metabólicas y moleculares agudas al RT realizado hasta el fallo no serían diferentes a las del entrenamiento de resistencia tradicional o entrenamiento aeróbico cuando la intensidad se controla adecuadamente. La función aguda del miocardio se mantendría, y tal vez aumentaría en respuesta al RT agudo de alta intensidad con un estímulo pequeño sobre la presión y sólo una mayor velocidad de contracción (es decir, frecuencia cardíaca y estímulo potencial a partir del volumen de sangre bombeado). La magnitud de la respuesta del flujo de sangre local agudo estaría determinada por la intensidad de la contracción. Sin embargo carecemos de datos obtenidos en humanos que demuestren el efecto sobre otros factores vasculares como la estimulación endotelial para la producción de óxido nítrico. La Figura 1 presenta un esquema de las respuestas agudas que se desencadenan al realizar RT hasta el fallo. Se necesitan más investigaciones en diferentes áreas para ayudar a evaluar el efecto del RT en las variables agudas. Sin embargo, una comprensión de las respuestas agudas frente al RT hasta el fallo nos permitiría identificar y discutir las adaptaciones crónicas que se producirían en la resistencia CV.
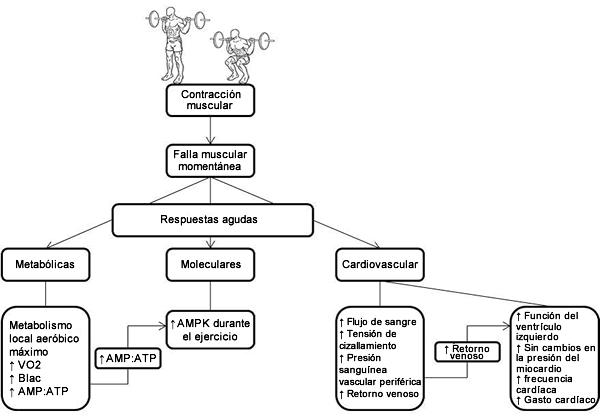
Figura 1. Esquema de las respuestas fisiológicas agudas al RT realizado hasta el fallo muscular momentánea.
5. PARTE 2: ADAPTACIONES CRÓNICAS AL RT
5.1 Adaptaciones Metabólicas y Moleculares
En la musculatura periférica se observan diferentes adaptaciones (entre las que se incluyen las enzimas oxidativas, fenotipo de las fibras musculares y la biogénesis mitocondrial) que pueden explicar la mejora en la resistencia CV en respuesta al RT. Se ha observado que el entrenamiento de la fuerza incrementa la concentración de ATP muscular en reposo (97) lo que sugiere que la producción de ATP aeróbica se incrementa. Sin embargo, este hallazgo no es una certeza absoluta proveniente exclusivamente de los resultados presentados por este estudio (97). Otra evidencia que examinó las intervenciones de RT demostró que se incrementan los marcadores de metabolismo aeróbico a través del RT. Esto apoyaría la conclusión de que se incrementa la producción de ATP mitocondrial. Por ejemplo, Jubrias et al. (79) realizaron una comparación entre un grupo que realizó RT, un grupo que realizó entrenamiento de resistencia tradicional y un grupo control. Los investigadores informaron un aumento en la capacidad de fosforilación oxidativa muscular máxima luego de la realización de RT (57%) en las personas mayores, y este aumento fue comparativamente mayor que el del grupo que realizó entrenamiento de la resistencia (31%). La intensidad del RT no fue controlada (10-15 repeticiones a 60-70% 1RM y luego 4-8 repeticiones a 70-85% 1RM con 3-5 sets) lo que, si bien podría haber involucrado un grado bastante alto de esfuerzo considerando los rangos de repetición con cargas similares (70, 133), no es consistente con la definición de esta revisión. Frontera y colegas (50) informaron una mayor actividad de enzimas mitocondriales (citrato sintasa) en los sujetos de edad avanzada luego de la realización de RT. Nuevamente, una definición específica de la intensidad del RT es imposible porque que los sujetos no realizaron RT hasta el fallo (3 series de 8 repeticiones a 80% de 1RM). Sin embargo, Tang et al. (145) observaron un aumento en el potencial oxidativo en varones desentrenados que realizaron RT hasta el fallo. Los autores observaron que el RT indujo una mejora significativa tanto en las enzimas de fosforilación de glucosa (hexoquinasa) como en las enzimas aeróbicas involucradas en el ciclo del ácido cítrico (citrato sintasa y ß-HAD).
Se ha sugerido que el RT podría favorecer la reducción de la densidad mitocondrial (86, 93, 95, 125, 149). Por consiguiente, sería contradictorio concluir que el RT aumenta la producción mitocondrial de ATP y la capacidad oxidativa. Sin embargo, solo uno de los estudios citados consideró el aumento concomitante en el volumen miofibrilar (es decir, hipertrofia), al medir la densidad mitocondrial. Luthi et al. (93) observaron que cuando se consideraba el aumento miofibrilar, el volumen mitocondrial (es decir, área transversal [CSA]) se mantenía sin cambios luego de RT. Notablemente, Jubrias et al. (79) informaron en su estudio que sólo el grupo de RT presentó un aumento en el volumen mitocondrial medido por espectroscopía de resonancia magnética, a pesar de también ser el único grupo que presentó un aumento en el CSA muscular. Así, parecería que el RT no afectaría el volumen mitocondrial cuando se considera la hipertrofia y podría aumentarlo. Dado que se ha demostrado que el AMPK induce la biogénesis mitocondrial (122, 157) lo que está claramente facilitado por el RT de alta intensidad, éste puede ser el factor responsable de la adaptación mitocondrial (incluyendo la regulación por incremento de las enzimas mitocondriales y de la proliferación mitocondrial).
El aumento en la capacidad oxidativa a través del RT podría deberse al cambio en el fenotipo de fibras musculares. Walker et al. (152) sugirieron que un aumento en potencia mecánica oxidativa por el RT podría deberse a un aumento en la actividad de fibras de tipo IIa. Un fenotipo de fibras de tipo IIa confiere una mayor capacidad oxidativa que las de tipo llx y esto explicaría la mejora en la capacidad aeróbica a través del RT (111). Staron et al. (137) observaron que los halterófilos tenían un porcentaje mayor de fibras de tipo lla en comparación con los controles, mientras que el grupo control tenía una proporción más alta de fibras de tipo llx. Esto puede parecer peculiar porque el principio del tamaño establecería que el RT de alta intensidad debería reclutar fibras de tipo llx y, por consiguiente, favorecer un aumento en las fibras (30). Sin embargo, los estudios de intervención confirman que el RT realizado hasta el fallo induce un cambio hacia un fenotipo de tipo lla (1, 57, 138, 155). Adams et al. (1) observaron una reducción significativa en fibras de tipo llx y un aumento concomitante en fibras de tipo lla, aunque no observaron cambios en el tipo I después de RT hasta el fallo. Staron et al. (138) demostraron una reducción similar en fibras de tipo llx y un aumento tanto en las de tipo lla como en las fibras de tipo I en el vasto lateral después de que los sujetos realizaran RT para los miembros inferiores hasta el fallo. Willoughby y Pelsue (155) observaron que 8 semanas de RT de alta intensidad indujeron regulación por incremento en la expresión de los genes de las fibras de tipo I y fibras de tipo lla y regulación hacia la baja (regulación negativa) de los genes de expresión de las fibras de tipo llx. Este cambio también se observó en las poblaciones de sujetos mayores (58, 87, 131).
Las adaptaciones que se producen por el RT se producirían, en parte, como resultado de la activación de AMPK cuando el entrenamiento se realiza en una intensidad alta (41, 51). La activación de AMPK explica el aumento observado en las enzimas mitocondriales y en la proliferación de las mismas. Además, el cambio en las fibras desde fenotipo tipo llx a fenotipo lla podría ser explicado porque las fibras de tipo llx presentan la mayor activación de AMPK (89). Así, no sería sorprendente que la mayor adaptación se produjera en ellas, porque la activación de AMPK en las fibras de tipo llx puede iniciar los cambios mitocondriales que conducen a un cambio hacia el fenotipo lla..
Las adaptaciones descriptas (regulación por incremento de las enzimas mitocondriales, mayor volumen mitocondrial y conversión a un fenotipo de fibras de tipo lla) han sido atribuidas a una mayor aptitud CV (109). Por consiguiente, es esperable que una adaptación en estos factores debida al RT también provoque un aumento concomitante en las variables de aptitud CV tal como se ha observado claramente.
5.2 Adaptaciones del miocardio
Es razonable esperar que el estímulo del miocardio y de la vasculatura periférica pueda impartir adaptaciones centrales al sistema de transporte de oxígeno y, por consiguiente, producir una mejora adicional a la aptitud CV. Tal como destacamos previamente se ha demostrado que el RT está asociado con un aumento en SBP (270 a 480 mmHg) (8, 94, 96), y se piensa que el aumento en la presión podría ser un estímulo para alterar el tamaño del ventrículo izquierdo (LV) (33). Si este es el caso, entonces, mejoraría los mecanismos de bombeo, el volumen cardíaco total y el gasto cardíaco máximo; y todos estos factores influyen en la aptitud CV (126, 139). Pero, como mencionamos en esta revisión, la presión puede no ser experimentada directamente por el propio corazón (90, 94).
Mientras que Pellicia et al. (115) informaron que los valores registrados en 100 atletas no superaban los límites superiores del rango normal, Baggish et al. (12) sugirieron que personas que habían realizado RT por un período de 90 días experimentaron hipertrofia del ventrículo izquierdo (LV). De manera similar, Pellicia et al. (115) observaron valores significativamente mayores en el espesor de la pared del LV en atletas de potencia que en controles sedentarios de la misma edad y valores similares de área superficial del cuerpo. Pluim et al. (120) en un meta análisis observaron una diferencia significativa entre el espesor de la pared del LV de atletas del grupo control (0,36 mm) y atletas entrenados en resistencia (0,39 mm) así como también entre atletas entrenados en resistencia (0,39 mm) y atletas entrenados en fuerza (0,44 mm). Es importante recordar que estos datos transversales no demuestran una relación causal. Las personas podrían haber escogido su deporte en base a su habilidad como resultado de diferencias cardíacas en lugar de presentar adaptaciones cardíacas crónicas como resultado de practicar su deporte.
Una revisión del efecto del RT en la hipertrofia cardíaca sugirió que RT no siempre produce cambios en la geometría del LV, entre los que se puede incluir el aumento en la hipertrofia del LV (64). Declarando que los datos están lejos de permitir la conclusión que el RT de cualquier tipo producirá estos cambios, Haykowsky et al. (64) observaron que 40% de todos los atletas entrenados en fuerza tienen una geometría de LV normal. Su trabajo de revisión consideró el tipo de RT (culturismo vs. rutinas de levantamiento de pesas) así como factores de edad y efectos de consumo de esteroides. Los autores sugirieron que las técnicas de levantamiento de pesas producirían los cambios más significativos en la masa del LV (hipertrofia concéntrica) mientras que el fisiculturismo produciría hipertrofia excéntrica. Lamentablemente, estos estudios no detallaron o controlaron las complejidades o la intensidad de los regímenes de entrenamiento (12, 115).
Entonces, en esencia las adaptaciones del miocardio a los diferentes factores de estrés provenientes del ejercicio sin consideración de la influencia genética o el uso de esteroides anabólicos son poco claras. Muchos autores declaran que la hipertrofia concéntrica del LV no es un producto del RT solo (151). De hecho, la literatura no aporta ninguna evidencia clara que sugiera que el RT de cualquier tipo presentará un estímulo suficiente para inducir adaptaciones estructurales del miocardio. Sin embargo, generalmente los estudios que han intentado examinar esta variable con respecto las variables del RT (sobre todo la intensidad) se controlan deficientemente. Las futuras investigaciones deberían examinar las adaptaciones del miocardio, e incluir el control del uso de esteroides.
5.3 Adaptaciones vasculares
Respecto a las adaptaciones vasculares que se proponen como causas del aumento en el flujo de sangre muscular local y del suministro de oxígeno, la tensión de cizallamiento es probablemente uno de los estímulos agudos para inducir adaptaciones vasculares. La investigación realizada por Tinken et al. (150), en donde además de examinar las respuestas agudas, también examinaron la adaptación al protocolo de RT de contracción del puño, informaron un aumento en la dilatación mediada por el flujo e hiperemia máxima reactiva de la arteria braquial (un índice de remodelación de las arterias de resistencia).
Habiendo observado que varones de edad avanzada que realizaban RT tenían un flujo de sangre y conductancia en la pierna completa significativamente mayores que los de los controles sedentarios, Miyachi et al. (103) sugirieron que el RT puede influir favorablemente en el flujo de sangre. Sin embargo, su estudio era una observación transversal. Es posible que las mejoras en el flujo de sangre y en la plasticidad vascular debidas a cualquier ejercicio estén limitadas a quienes tienen edad avanzada o padecen una enfermedad vascular, porque la mayoría de las investigaciones se han enfocado en estas poblaciones. Por otro lado, Tanimoto et al. (146) observaron un aumento significativo en el flujo de sangre femoral en varones jóvenes saludables después de 13 semanas de RT hasta el fallo. Ellos usaron dos grupos, uno que realizaba ejercicio con una carga baja y con repeticiones de larga duración (Grupo LST; ~55% a 60% 1 RM, 3 seg fase concéntrica y 3 seg fase excéntrica) y el otro que utilizó una carga alta y repeticiones de corta duración (Grupo HN; ~85% a 90% 1RM, 1 seg fase excéntrica, 1 seg fase concéntrica).Los grupos LST y HN aumentaron significativamente el flujo de sangre femoral (18% y 35%, respectivamente) y no se observaron diferencias significativas entre los grupos. Podría haber existido un efecto de carga debido al mayor aumento en el grupo HN, pero esto no quedo claro e investigaciones futuras que intenten reproducir este resultado ayudarían a aclarar si efectivamente existe una diferencia. Está claro que el RT hasta el fallo produce adaptaciones vasculares en los varones jóvenes saludables. En apoyo a las conclusiones efectuadas por Miyachi et al. (103), y a los resultados obtenidos por Tanimoto et al. (146), Anton et al. (4) observaron una mejora significativa en el flujo de sangre femoral (55% a 60%) en varones y mujeres de edad avanzada después de 13 semanas de RT hasta el fallo. Estos cambios fueron independientes de los cambios en los vasoconstrictores como la endotelina-1 y angiotensina-ll, y no se observó ningún cambio en el espesor de la pared arterial. Rakobowchuk et al. (121) observaron una mejor función endotelial a través de un mayor flujo máximo y de un mayor diámetro de la arteria braquial independientemente de la dilatación mediada por el flujo después de RT hasta el fallo. Estos resultados parecen apoyar la noción que una mejor función vascular puede ser predominantemente un resultado de adaptación a nivel arteriolar.
Brown (24) estableció que la proliferación de capilares y 'la arteriolización” representan una respuesta integral a la remodelación vascular coronaria, y lo mismo puede ser aplicable a la remodelación vascular periférica. Green et al. (57) observaron un aumento en los contactos capilares como resultado de 12 semanas de RT hasta el fallo. Adicionalmente, Hepple y colegas (67) demostraron que varones de edad avanzada que realizaron RT hasta el fallo durante un período de 9 semanas aumentaron significativamente los contactos capilares y la relación entre capilar y fibra. Si solo se realizaba entrenamiento aeróbico, los mismos cambios tardaban 18 semanas en alcanzar significancia. La intensidad de entrenamiento aeróbico no se informó, pero parecería razonable sugerir que habría sido probable significativamente menor que la intensidad del RT, aunque también debe haber sido realizado hasta el agotamiento. Dado que la única medida cuantificable de intensidad del ejercicio es el fallo muscular momentáneo y generalmente el entrenamiento aeróbico tradicional no se realiza de esta manera, entonces, esto podría explicar las adaptaciones más rápidas en el grupo que realizó RT. Harris (61) comentó que el ejercicio de resistencia afecta positivamente los cambios capilares, pero hay datos contradictorios sobre el entrenamiento de la fuerza (RT). También destacó que la intensidad del entrenamiento podría ser un factor importante, algo que ha sido respaldado por los datos obtenidos por Hepple et al. (67).
También debemos destacar que los estudios han informado que el RT de alta intensidad no mejora la densidad capilar (67, 153). Sin embargo, los dos estudios citados aquí observaron un aumento en el CSA del músculo después del entrenamiento. De la misma manera que los aumentos de CSA confunden las mediciones de densidad mitocondrial, es probable que afecten las mediciones de densidad capilar. En contraste, Hagerman et al. (2000) observaron tanto un aumento en el CSA muscular como un aumento en la capilarización después de la realización de RT hasta el fallo. Los datos de Hepple et al. (67) y Green et al. (57), que identificaron los contactos capilares y la relación entre capilar y fibra, son probablemente más indicativos de la remodelación vascular en respuesta al RT (porque las mediciones no están afectadas por los aumentos concomitantes en el CSA). Entonces, parecería que se produce una remodelación vascular significativa a nivel muscular local periférico como resultado del RT hasta el fallo que puede aumentar el suministro de oxígeno local.
6. RESUMEN DE LA PARTE 2: ADAPTACIONES CRÓNICAS AL RT
Muchas investigaciones han demostrado la existencia de adaptaciones fisiológicas positivas que pueden mediar la mejora observada en la aptitud CV como resultado del RT. También está claro que estas adaptaciones son, en su mayor parte, un resultado de RT de alta intensidad (es decir, realizado hasta el fallo). Las adaptaciones que se producen, regulación por incremento de las enzimas mitocondriales, proliferación mitocondrial y conversión hacia un fenotipo de fibras de tipo lla pueden ser las adaptaciones fisiológicas que sustentan la mejora observada en las variables de aptitud CV. Hay poca evidencia que sustente la afirmación que el miocardio es estimulado directamente, y que cualquier adaptación subsecuente se promueve dentro de él como respuesta al RT de alta intensidad y parece improbable que éste sea un mecanismo involucrado en la mejora de la aptitud CV como resultado del RT. Sin embargo, se han observado adaptaciones vasculares, entre las que se incluye la capilarización, dentro de la musculatura periférica como respuesta al RT hasta el fallo. Es probable que estas adaptaciones contribuyan con la mejora en la aptitud CV a través de un aumento en el suministro local de oxígeno muscular. En la Figura 2 se muestra un esquema de las adaptaciones crónicas producidas por el RT realizado hasta el fallo. Parecería que tales adaptaciones no dependen de una modalidad de ejercicio particular porque se han observado adaptaciones similares por medio de otras formas de entrenamiento de alta intensidad y los estudios comparativos de diferentes modalidades no favorecen claramente una forma por encima de otra. Nosotros sugerimos que el factor clave para determinar las adaptaciones fisiológicas que promueven la aptitud CV es la contracción muscular intensa.
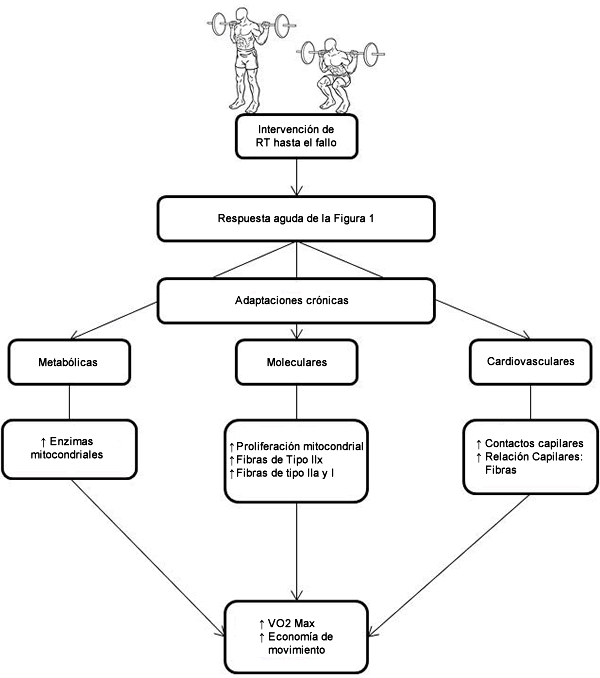
Figura 2. Esquema de las adaptaciones fisiológicas crónicas y mejoras en CV a partir del RT realizado hasta el fallo muscular momentánea.
7. CONCLUSIONES
7.1 Áreas para las investigaciones futuras
En esta revisión hemos destacado cuales son las áreas en las cuales las investigaciones están inconclusas o ausentes. Las áreas que deben ser investigadas en el futuro incluyen: (a) Las mejoras en Tlac sólo fueron evidenciadas en poblaciones de sujetos desentrenados en un estudio por lo que se necesitan estudios adicionales para esclarecer esta respuesta; (b) Si bien la respuesta del VO2 del cuerpo entero frente al ejercicio tiene poco impacto en las adaptaciones CV, estudios adicionales que analicen el VO2 durante RT hasta el fallo (sin obtener promedios que incluyan los periodos sin ejercicio) podrían ayudar a determinar el papel del VO2 del músculo activo local en las adaptaciones CV; (c) Además, considerado el papel de la intensidad y que las repeticiones de mayor duración pueden tener una intensidad mayor que las de menor duración, también sería interesante examinar el efecto de esta variable en el VO2 durante RT; (d) también sería importante realizar investigaciones adicionales para identificar el estímulo responsable de la adaptación vascular (es decir, capilarización), así como también establecer el rol de la producción endotelial de oxido nítrico y la participación vascular en la respuesta al RT; y (e) En referencia a las adaptaciones CV centrales, sería útil realizar estudios que analicen las intervenciones de RT controladas en poblaciones controladas para el uso de esteroides para aclarar si el RT realizado hasta el fallo produce adaptaciones en el miocardio.
7.2 Aplicaciones prácticas
En base a esta revisión, debemos destacar varias aplicaciones prácticas. Si bien se ha demostrado que las adaptaciones fisiológicas que promueven la aptitud CV son similares en las modalidades de ejercicio de resistencia aeróbico tradicional y ejercicios de entrenamiento de la fuerza (RT), los estudios comparativos que han controlado la intensidad de RT no sugieren ninguna diferencia significativa. La magnitud de las respuestas fisiológicas puede ser diferente en las diferentes poblaciones. Por ejemplo, los atletas entrenados muestran una mejora pequeña en el VO2max en respuesta al RT lo que probablemente se deba a su nivel de entrenamiento. Por consiguiente, el valor de RT puede diferir con respecto a la población considerada. Es muy probable que las personas que no están entrenadas o que no participan en competencias deportivas organizadas, pero que tienen deseos de mejorar su aptitud CV puedan obtener beneficios del RT realizado hasta el fallo. De hecho, esta revisión sugiere que el RT hasta el fallo puede producir efectos de aptitud CV y provocar simultáneamente mejoras en la fuerza, potencia y otras variables relacionadas a la salud y a la aptitud física (48). Esto sería una inversión eficaz de tiempo porque la persona no tendría que realizar varios programas de entrenamiento independientes para diferentes aspectos de la aptitud física. Al contrario, el valor del RT para mejorar la CV de un atleta y potencialmente el rendimiento deportivo sería menor. Aunque las adaptaciones fisiológicas producidas serían similares a las que se obtienen si el atleta debe realizar RT, la participación en otras modalidades de entrenamiento puede ser esencial para asegurar que las habilidades motoras específicas y otras variables involucradas con su rendimiento también sean entrenadas. Está más allá del alcance de esta revisión sugerir otras maneras optimas para utilizar el RT (es decir, carga, volumen establecido, y/o frecuencia) para mejorar la aptitud CV porque no hay ningún estudio publicado sobre este tema. Por consiguiente, para mejorar la aptitud CV el RT se recomienda para atletas y no atletas, ya sea de manera tradicional o en forma de circuito, y recomendamos que se realice hasta el fallo tal como lo sostienen Fisher et al. (48).
7.3 CONCLUSION
El entrenamiento de la fuerza realizado hasta el fallo puede inducir efectos fisiológicos agudos y crónicos que parecen ser similares a los del entrenamiento de resistencia aeróbico, que su vez producen mejoras similares en la aptitud CV. Se ha realizado un número limitado de estudios de comparación directa entre las dos modalidades. Los estudios que fueron controlados apropiadamente sugieren que no existirían diferencias significativas en la aptitud CV o en las respuestas fisiológicas. Esta revisión también ha resaltado cuales serían las áreas dónde la investigación actual está inconclusa. Por consiguiente, es importante que se realicen investigaciones para comprender de qué manera el RT hasta el fallo permite lograr una mayor aptitud CV. Nosotros somos optimistas y confiamos que este trabajo de revisión ayudará a promover un cambio de paradigma en esta área de entrenamiento e investigación. Es erróneo identificar una modalidad particular de ejercicio como “aeróbica” o “CV”. La magnitud en que cualquier modalidad de ejercicio produce adaptaciones de aptitud CV parece depender principalmente de la intensidad del ejercicio. Las adaptaciones crónicas parecen residir predominantemente en el nivel muscular y vascular periférico. De hecho, nosotros no somos los primeros en considerar esta falsa dicotomía entre RT y entrenamiento aeróbico (119). Phillips y Winnett (119) cuestionaron la dicotomía de RT/aeróbico con respecto a los resultados de salud. Ellos, también, sugieren que es importante reconsiderar el valor del RT. Esperamos que esta revisión anime a investigadores y profesionales a considerar las implicaciones en el diseño e interpretación de las futuras investigaciones así como al prescribir los ejercicios con el propósito de mejorar la aptitud CV. Así, en conclusión, sostenemos que la realización de RT hasta el fallo producirá una mejora significativa en la aptitud CV a través de adaptaciones fisiológicas como la regulación por incremento de las enzimas mitocondriales, proliferación mitocondrial, la conversión hacia un fenotipo de fibras de tipo lla y capilarización.
Dirección para correspondencia: Centre for Health, Exercise and Sport Science, Southampton Solent University, Southampton, Hampshire, United Kingdom, S014 OYN. Phone (02380319606); Email.James.Steele@Solent.ac.uk
Referencias
1. Adams G.R., Hather B.M., Baldwin K.M., et al. (1993). Skeletal muscle myosin heavy chain composition and resistance training. J. Appl. Physiol. 74(2):911-915.
2. Amirfallah N. (2003). Die Auswirkungen dynamischen Krafttrainings nach dem Nautilus-Prinzip auf kardiozirkulatorische Parameter und Ausdauerleistungsfahigkeit (The effects of resistance training according to the Nautilus principles on cardiocirculatory parameters and endurance). Angenommen vom Fachbereich Humanmedizin der Philipps-Universitat Marburg.
3. Andersen P., Saltin B. (1985). Maximal perfusión of skeletal muscle in man. J Physiol.366(1): 233-249.
4. Anton M.M., Cortez-Cooper M.Y., DeVan A.E. et al. (2006). Resistance training increases basal limb blood flow and vascular conductance in aging humans. J. Appl. Physiol. 101 (5): 1351-1355.
5. Arany Z., Lebrasseur N., Morris C. et al. (2007). The transcriptional coactivator PGC-1 [beta] drives the formation of oxidative type MX fibers in skeletal muscle. Cell. Metab.5(1):35-46.
6. Arent S.M., Landers D.M., Matt K.S., et al. (2005). Dose-response and mechanistic issues in the resistance training and affect relationship. J. Sport. Exerc. Psych.27:92-110.
7. Asmussen E. (1953). Positive and negative muscular work. Acta Physiol. Scand.28(4):364-382.
8. Astorino T.A., Rohmann R.L., Firth K., et al. (2007). Caffeine-induced changes in cardiovascular function during resistance training. Int. J. Sport Nutr. Exerc. Metab.17(5):468-477.
9. Astrand P., Rodahl K., Dahl H.A. and Stramme S.B. (2003). Textbook of Work Physiology: Physiological Basis of Exercise. (4th Edition). Champaign, United States: Human Kinetics.
10. Atherton P.J., Babraj J., Smith K., et al. (2005). Selective activation of AMPK-PGC-1a or PKB-TSC2-mTOR signaling can explain specific adaptive responses to endurance or resistance training like electrical muscle stimulation. FASEBJ.19(7):786-789.
11. Baar K. (2006). Training for endurance and strength: lessons from cell signaling. Med. Sci. Sports Exerc.38(11): 1939-1944.
12. Baggish A.L., Wang F., Weiner R.B. et al. (2008). Training-specific changes in cardiac structure and function: a prospective and longitudinal assessment of competitive athletes. J. Appl. Physiol. 104(4): 1121-1128.
13. Barden J., Lawrenson L., Poole J.G. et al. (2007). Limitations to vasodilatory capacity and V02 max in trained human skeletal muscle. Am. J. Physiol. Heart Circ. Physiol.292(5):H2491-2497.
14. Barreto A.C., Maior A.S., Pedro M., et al. (2010). Effect of different resistance exercise repetition velocities on excess post-exercise oxygen consumption and energetic expenditure. International Sport Med. Journal..
15. Bassett D.R., Howley E.T. (1997). Maximal oxygen uptake: 'classical' versus 'contemporary' viewpoints. Med Sci Sports Exerc.29(5):591-603.
16. Bassett D.R., Howley E.T. (2000). Limiting factors for maximum oxygen uptake and determinants of endurance performance. Med. Sci. Sports Exerc.32(1 ):70-84.
17. Beckham S., Earnest C. (2000). Metabolic cost of free weight circuit weight training. J. Sports Med. Phys. Fitness. 40(2): 118-125.
18. Benito-Peinado P.J., Álvarez-Sánchez M., Diaz-Molina V., et al. (2010). Aerobic energy expenditure and intensity prediction during a specific circuit weight training: A pilot study. International Sport Med. Journal.
19. Bishop D., Jenkins D.G., Mackinnon L.T., et al. (1999). The effects of strength training on endurance performance and muscle characteristics. Med. Sci. Sports Exerc.31 (6):886-891.
20. Bloomer R.J. (2005). Energy cost of moderate-duration resistance and aerobic exercise. J. Strength Cond. Res. 19(4): 878-882.
21. Bolster D.R., Kubica N., Crazier S.J. et al. (2003). Immediate response of mammalian target of rampamycin (mTOR)-mediated signalling following acute resistance exercise in rat skeletal muscle. J. Physiol.553 (1):213-220.
22. Bolster D.R., Jefferson L.S., Kimball S.R. (2004). Regulation of protein synthesis associated with skeletal muscle hypertrophy by insulin-, amino acid- and exercise-induced signalling. Proc. Nutr. Soc.63 (2):351-356.
23. Brinkley T.E., Fenty-Stewart N.M., Park J.Y. et al. (2009). Plasma Nitrate/Nitrite Levels are Unchanged after Long-TermAerobic Exercise Training in Older Adults. Nitric Oxide.21(3-4):234-238.
24. Brown M.D. (2003). Exercise and coronary vascular remodelling in the healthy heart. Exp. Physiol.88 (5):645-658.
25. Bruce-Low S., Smith D. (2007). Explosive exercise in sports training: A critical review. JEPonline.10 (1):21-33.
26. Buda A.J., Pinsky M.R., Ingels Jnr N.B., et al. (1979). Effect of intrathoracic pressure on left ventricular performance. N. Engl. J. Med.301:453-459.
27. Burgomaster K.A., Hughes S.C., Heigenhauser G.J.F., et al. (2005). Six sessions of sprint interval training increases muscle oxidative potential and cycle endurance capacity in humans. J. Appl. Physiol.98:1985-1990.
28. Burleson M.A., O'Bryant H.S., Stone M.H. et al. (1998). Effect of weight training exercise and treadmill exercise on post-exercise oxygen consumption. Med. Sci. Sports Exerc.30 (4):518-522.
29. Calbet J.A., Lundby C., Sander M., et al. (2006). Effects of ATP-induced leg vasodilation on V02 peak and leg 02 extraction during maximal exercise in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 291 (2):R447-R453.
30. Carpinelli R. (2008). The size principle and a critical analysis of the unsubstantiated Heavier-is-better recommendation for resistance training. J. Exerc. Sci. Fit.6 (2):67-86 .
31. Charro M.A., Aoki M.S., Coutts A.J., et al. (2010). Hormonal metabolic and perceptual responses to different resistance training systems. J. Sports Med. Phys. Fitness.50:229-234.
32. Coffey V.G., Hawley J.A. (2006). Training for performance: Insights from molecular biology. IntJ Sports Physiol Perform. 1(3): 284-292.
33. Colan S.D. (1992). Mechanics of left ventricular systolic and diastolic function in physiologic hypertrophy of the athlete heart. Cardiol. Clin. 10:227-240.
34. Colliander E.B., Tesch P.A. (1988). Blood pressure in resistance trained athletes. Can. J. Appl. Sport Sci.13:31-34.
35. Collins M.A., Cureton K.J., Hill D.W. et al. (1991). Relationship of heart rate to oxygen uptake during weight lifting exercise. Med. Sci. Sports Exerc. 23(5):636-640.
36. Collins J.A., Fauser C.J.M. (2005). Balancing the strengths of systematic and narrative reviews. Hum. Reprod. Update. 11 (2): 103-104.
37. Cooper K. (1970). The New Aerobics. New York: Bantam Books.
38. Dalleck L., Bushman T.T., Crain R.D., et al. (2010). Dose-response relationship between interval training frequency and magnitude of improvement in lactate threshold. Int. J. Sports Med.31 (8): 567-571.
39. Dentón J., Cronin J.B. (2006). Kinematic, kinetic, and blood lactate profiles of continuous and intraset rest loading schemes. J. Strength Cond. Res. 20 (3):528-534.
40. DeVan A.E., Anton M.M., Cook J.N. et al. (2005). Acute effects of resistance training on arterial compliance. J. Appl. Physiol.98:2287-2291.
41. Dreyer H.C., Fujita S., Cadenas J.G., et al. (2006). Resistance exercise increase AMPK activity and reduces 4E-BP1 phosphorylation and protein synthesis in human skeletal muscle. J. Physiol.576 (2):613-624.
42. Dubouchaud H., Butterfield G.E., Wolfel E.E., et al. (2000). Endurance training, expression, and physiology of LDH, MCT1 and MCT4 in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab.278:E572- 579.
43. Dudley G.A. (1988). Metabolic consequences of resistive-type exercise. Med. Sci. Sports Exerc. 20(5):S158-161.
44. Dunker D.J., Bache R.J. (2008). Regulation of coronary blood flow during exercise. Physiol. Rev. 88:1009-1086.
45. Ebben W.P., Kindler A.G., Chirdon K.A., et al. (2004). The effect of high-load vs. high-repetition training on endurance performance. J Strength Cond Res.18:513-517.
46. Ekiund B., Kaijser L. (1974). Forearm blood flow after isometric contraction at different loads in relation to potentially vasodilating substances. Scand. J. Clin. Lab. Invest. 34 (1):23-29.
47. Enoksen E., Shalfawi S.A.I., Tonnessen E. (2011). The Effect of High-vs Low-Intensity Training on Aerobic Capacity in Well-Trained Male Middle-Distance Runners. J. Strength Cond. Res.25(3):812-818.
48. Fisher J., Steele J., Bruce-Low S., Smith D. (2011). Evidence based resistance training recommendations. Medicina Sportiva.15(3): 147-162.
49. Foppa M. Duncan B.B., and Rohde L.E.P. (2005). Echocardiography-based left ventricular mass estimation. How should we define hypertrophy? Cardiovasc. Ultrasound.3 (17): 1-13.
50. Frontera W.R., Meredith C.N., O'Reilly K.P. et al. (1990). Strength determinants of V02max in older men. J. Appl. Physiol. 68(1 ):329-333.
51. Frosig C., Jorgensen S.B., Hardie D.G. et al. (2004). 5'-AMP-activated protein kinase activity and protein expression are regulated by endurance training in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab.286 (3): E411 -417.
52. Gentil P., Oliveira E., Bottaro M. (2006). Time under tension and blood lactate response during four different resistance training methods. J. Physiol. Anthropol. 25 (5): 339-344.
53. Gibala M.J., Little J.P. van Essen M., et al. (2006). Short-term sprint interval training versus traditional endurance training: similar initial adaptations in human skeletal muscle and exercise performance. J. Physiol.575 (3):901-911.
54. Gibala M.J., McGee S.L., Garnham A.P., et al. (2009). Brief intense interval exercise activates AMPK and p38 MAPK signalling and increase the expression of PGC-1a in human skeletal muscle. J. Appl. Physiol.; 106:929-934.
55. Goldberg L., Elliot D.L., Kuehl K. S. (1994). A comparison of the cardiovascular effects of running and weight training. J Strength Cond Res.8:219-224.
56. Gotshalk L.A., Berger R.A., Kramer W.J. (2004). Cardiovascular responses to a high-volume continuous circuit resistance training protocol. J. Strength Cond. Res.18 (4):760-764.
57. Green H., Goreham C., Ouyang J., et al. (1998). Regulation of fiber size, oxidative potential, and capillarization in human muscle by resistance exercise. Am. J. Physiol.276:R591-596.
58. Hagerman F.C., Walsh S.J., Staron R.S., et al. (2000). Effects of high-intensity resistance training on untrained older men. I. Strength, cardiovascular, and metabolic responses. J. Gerontol. A. Biol. Sci. Med. Sci.55 (7): B336-346.
59. Hardie D.G., Salt I.P., Hawkley S.A., et al. (1999). AMP-activated protein kinase: and ultrasensitive system for monitoring cellular energy change. Biochem. J. 338(3):717-722.
60. Hardie D.G., Sakamoto K. (2006). AMPK: A key sensor of fuel and energy status in skeletal muscle. Physiology.21:48-60.
61. Harris B.A. (2005). The influence of endurance and resistance exercise on muscle capillarisation in the elderly: a review. Acta Physiol. Scand.185:89-87.
62. Hartman M.J., Fields D.A., Byrne I.S.M., et al. (2007). Resistance training improves metabolic economy during functional tasks in older adults. J. Strength Cond. Res.21:91-5.
63. Haykowsky M., Taylor D., Teo K., et al. (2001). Left ventricular wall stress during leg press exercise performed with a brief valsalva maneuver. Chest.119:150-154.
64. Haykowsky M.J., Dressendorfer R., Taylor D, et al. (2002). Resistance training and cardiac hypertrophy; unravelling the training effect. Sports Med. 32 (13):837-849.
65. Haykowsky M.J., Teo K.K., Quinney A.H., et al. (2000). Effects of long term resistance training on left ventricular morphology. Can. J. Cardiol. 16(1):35-38.
66. Helgerud J., Hoydal K., Wang E., et al. (2007). Aerobic high-intensity intervals improve V02max more than moderate training. Med. Sci. Sports Exerc.39 (4):665-671.
67. Hepple R.T., Mackinnon S.L.M., Goodman J.M., et al. (1997). Resistance and aerobic training in older men: effects on V02peak and the capillary supply to skeletal muscle. J. Appl. Physiol. 82:1305-1310.
68. Hermansen L. (1973). Oxygen transport during exercise in human subjects. Acta. Physiol. Scand. Suppl.399:1-104.
69. Hickson R.C., Rosenkoetter M.A., Brown M.M. (1980). Strength training effects on aerobic power and short term endurance. Med. Sci. Sports Exerc.12:336-339.
70. Hoeger W.W.K., Hopkins D.R., Barette S.L., et al. (1990). Relationship between repetitions and selected percentages of one repetition maximum: a comparison between untrained and trained males and females. J. Strength Cond. Res.4:46-54.
71. Huggett D.L., Elliott I.D., Overend T.J., et al. (2004). Comparison of heart rate and blood-pressure increases during isokinetic eccentric versus isometric exercise in older adults. J. Aging Phys. Act.11:157-169.
72. Hultman E., Sjoholm H. (1983). Energy metabolism and contraction force of human skeletal muscle In Situ during electrical stimulation. J. Physiol.345:525-532.
73. Hunter G.R., Seelhorst D., Snyder S. (2003). Comparison of metabolic and heart rate responses to super slow vs. traditional resistance training. J. Strength Cond. Res. 17(1):76-81.
74. Hurley B.F., Seals D.R., Ehsani A.A., et al. (1984). Effects of high-intensity strength training on cardiovascular function. Med. Sci. Sports Exerc. 16 (5):483-488.
75. Johnston R.E., Quinn T.J., Kertzer R., et al. (1997). Strength training in female distance runners: impact on running economy. J Strength Cond Res.11:224-229.
76. Jones A.M., Carter H. (2000). The effect of endurance training on parameters of aerobic fitness. Sports Med. 29:373-86.
77. Jones A.M., Doust J.H. (2001). Limitations to submaximal exercise performance In: Eston R, Reilly T (Editors). Kinanthropometry and Exercise Physiology Laboratory Manual: Tests, Procedures and Data. Volume 2: Exercise Physiology. New York: Routledge,: 235-263.
78. Komi P.V. (1986). The stretch shortening cycle and human power output, in Jones N. L., McCartney N., McComas A.J., (Editors). Human Muscle Power. Champaign: Human Kinetics. 27-40.
79. Jubrias S.A., Esselman P.C., Price L..B, et al. (2001). Large energetic adaptations of elderly muscle to resistance and endurance training. J. Appl. Physiol.90:1663-1670.
80. Jung A.P. (2003). The impact of resistance training on distance running performance. Sports Med.33(7):539-552.
81. Jurimae T., Jurimae J., Pihl E. (2000). Circulatory response to single circuit weight and walking training sessions of similar energy cost in middle aged overweight females. Clin. Physiol.20(2): 143-149.
82. Karlsdottir A.E., Foster C., Porcari J.P., et al. (2002). Hemodynamic responses during aerobic and resistance exercise. J. Cardiopulm. Rehabil. 22:170-177.
83. Kendall T.L., Black C.D., Elder C.P., et al. (2006). Determining the extent of nueral activation during maximal effort. Med. Sci. Sports Exerc. 38:1470-1475.
84. Koopman R., Zorenc A.H.G., Gransier R.J.J., et al. (2006). Increase in S6K1 phosphorylation in human skeletal muscle following resistance exercise occurs mainly in type II muscle fibers. Am. J. Physiol. Endocrinol. Metab.290(6):E1245-1252.
85. Kraemer W.J., Spiering B.A. (2006). Skeletal muscle physiology: Plasticity and responses to exercise. Horm. Res.66(supp 1):2-16.
86. Kraemer W.J., Deschenes M.R., Fleck S.J. (1988). Physiological adaptations to resistance exercise: implications for athletic conditioning. Sports Med.6:246-256.
87. Kryger A.L, Andersen J.L. (2007). Resistance training in the oldest old: consequences for muscle strength, fiber size, and MHC isoforms. Scand. J. Med. Sci. Sports.17:422-430.
88. Laursen P.B., Shing C.M., Peake J.M., et al. (2005). Influence of high-intensity interval training on adaptations in well trained cyclists. J. Strength Cond. Res. 19 (3):527-533.
89. Lee-Young R.S., Canny B.J., Myers D.E., et al. (2008). AMPK activation is fiber type specific in human skeletal muscle: effects of exercise and short-term exercise training. J. Appl. Physiol.107:283-289.
90. Lentini A.C., McKelvie R.S., McCartney N. et al. (1993). Left ventricular response in healthy young men during heavy-intensity weight-lifting exercise. J. Appl. Physiol.75:2703-2710.
91. Lin J., Wu H., Tarr P.T., et al. (2002). Transcriptional co-activator PGC-1 alpha drives the formation of slow-twitch muscle fibers. Nature.418 (6899):797-801.
92. Mayer L.B., Stifter G., Putz S., et al. (1993). Effects of dichloroacetate on exercise performance in healthy volunteers. Phlugers Arch.423 (3-4):251-254.
93. Luthi J.M., Howald H., Claassen H., et al. (1986). Structural changes in skeletal muscle tissue with heavy resistance exercise. Int. J. Sports Med.7(3):123-127.
94. MacDougall J.D., McKelvie R.S., Moroz D.E., et al. (1992). Factors affecting blood pressure during heavy weight lifting and static contractions. J. Apply. Physiol.73:1590-1597.
95. MacDougal J.D., Sale D.G., Moroz J.R., et al. (1979). Mitochondrial volume density in human skeletal muscle following heavy resistance training. Med. Sci. Sports Exerc.11:164-166.
96. MacDougall J.D., Tuxen D., Sale D.G., et al. (1985). Arterial blood pressure response to heavy resistance exercise. J. Appl. Physiol.58:785-790.
97. MacDougall J.D., Ward G.R., Sale DG, et al. (1977). Biochemical adaptation of human skeletal muscle to heavy resistance training and immobilization. J. Appl. Physiol.43(3):700-703.
98. Marcinik E.J., Potts J., Schlaback G., et al. (1991). Effects of strength training on lactate threshold and endurance performance. Med. Sci. Sports Exerc. 23:739-743.
99. Messier S.P., Dill M.E. (1985). Alterations in strength and maximum oxygen consumption consequent to nautilus circuit weight training. Res. Q. Exerc. Sport.56:345-351.
100. Meyer K., Hajric R., Westbrook S. et al. (1999). Hemodynamic responses during leg press exercise in patients with chronic congestive heart failure. Am. J. Cardiol.83(11): 1537-1543.
101. Midgley A.W., McNaughton L.R., Wilkinson M. (2006). Is there an optimal training intensity for enhancing the maximal oxygen uptake of distance runners? Empirical research findings, current opinions, physiological rationale and practical recommendations. Sports Med.36(2): 117-132.
102. Millet G.P., Jaouen B.J., Borrani F. et al. (2002). Effects of concurrent endurance and strength training on running economy and V02 kinetics. Med Sci Sports Exerc.34(8): 1351-1359.
103. Miyachi M., Tanaka H., Kawano H. et al. (2005). Lack of age related decreases in basal whole leg blood flow in resistance trained men. JAppI Physiol.99:1384-1390.
104. Morganroth J., Maron B.J., Henry W.L., et al. (1975). Comparative Left Ventricular dimensions in trained athletes. Ann. Intern. Med.82:521-524.
105. Nader G.A. (2006). Concurrent strength and endurance training: From molecules to man. Med Sci Sports Exerc.38 (11): 1965-1970.
106. Noakes T.D. (1988). Implications of exercise testing for prediction of athletic performance: a contemporary perspective. Med. Sci. Sports Exerc.20:319-330.
107. Noakes T.D. (1997). Challenging beliefs: ex Africa simper aliquid novi. Med. Sci. Sports Exer.29(5):571-590.
108. Noakes T.D. (1998). Maximal oxygen uptake: 'classical' versus 'contemporary' viewpoints: A rebuttal. Med. Sci. Sports Exerc. 30 (9): 1381 -1398.
109. Noakes T.D. (2003). Lore of Running. (4th Edition). Champaign: Human Kinetics.
110. Noakes T.D, St Clair Gibson A. (2004). Logical limitations to the "catastrophe" models of fatigue during exercise in humans. Br. J. Sports Med.38 (5):648-649.
111. O'Hara R., Khan M., Pohlman R. and Schlub J. (2004). Leg resistance training: Effects upon V02peak and skeletal muscle myoplasticity. J Exerc Physiol.7(5):27-43.
112. O'Hara R.B., Schlub J.F., Siejack T.R. et al. (2004). Increased volume resistance training: effects upon predicted aerobic fitness in a select group of air force men. ASCM'S Health & Fitness Journal.8:4-8.
113. Okamoto T., Masuhara M., Ikuta K. (2006). Cardiovascular responses induced during high intensity eccentric and concentric isokinetic muscle contraction in healthy young adults. Clin. Physiol. Funct. Imaging. 26 (1):39-44.
114. Paavolainen L., Hakkinen K., Hamalainen I. et al. (1999). Explosive-strength training improves 5-km running time by improving running economy and muscle power. J. Appl. Physiol.86: 1527-1533.
115. Pellicia A., Spataro A., Caselli G. et al. (1993). Absence of left ventricular wall thickening in athletes in engaged in intense power training. Am. J. Cardiol. 72:1048-1054.
116. Peterson J.A. (1975). Total conditioning: A case study. Athletic. Journal.56:40-55.
117. Phillips W.T., Ziuraitis J.R. (2003). Energy cost of the ACSM single-set resistance training protocol. J. Strength Cond. Res.17(2): 350-355.
118. Phillips W.T., Ziuraitis J.R. (2004). Energy cost of the single-set resistance training in older adults. J. Strength Cond Res. 18 (3):606-609.
119. Phillips S.M., Winett. (2010). Uncomplicated resistance training and health related outcomes: Evidence for a public health mandate. Curr. Sports Med. Rep. 9 (4):208-213.
120. Pluim B.M., Zwinderman A.H., van der Laarse A., et al. (2000). The athlete's heart: A meta-analysis of cardiac structure and function. Circulation.100:336-344.
121. Rakobowchhuk M., McGowan C.L., de Groot P.C. et al. (2005). Endothelial function of young healthy males following whole body resistance training. J. Appl. Physiol.98:2185-2190.
122. Reznick R.M., Shuman G.L. (2006). The role of AMP-activated protein kinase in mitochondrial biogenesis. J. Physiol.574(1):33-39.
123. Robergs R.A. (2000). An exercise physiologists 'contemporary' interpretations of the 'ugly and creaking edifices' of the V02 max concept. JEPonline.4(1):1- 44.
124. Robergs R.A., Gordon T., Reynolds J., et al. (2007). ;Energy expenditure during bench press and squat exercises. J. Strength Cond. Res. 21(1): 123-130.
125. Sale D.G., Jacobs I., MacDougall J.D., et al. (1990). Comparison of two regimens of concurrent strength and endurance training. Med. Sci. Sports Exerc.22 (3):348-356.
126. Saltin B., Calbet J.A.L. Point: (2006). In health and in a normoxic environment V02 max is limited primarily by cardiac output and locomotor muscle blood flow. J. Appl. Physiol.100 (1): 744-748.
127. Salway J.G. (2004). Metabolism at a Glance Oxford: Blackwell Publishing.
128. Sargeant A.J. (2007). Structural and functional determinants of muscle power. Exp. Physiol.92 (2):323-331.
129. Saunders P.U., Telford R.D., Pyne D.B., et al. (2006). Short-term plyometric training improves running economy in highly trained middle and long distance runners. J. Strength Cond. Res. 20: 947-954.
130. Schlub J.F., Siejack T.R., O'Hara R.B., et al. (2003). Effects of a traditional vs. non-traditional physical training program on submaximal cycle ergometry scores in USAF men. Aviat. Space Environ. Med.74:406-411.
131. Sharman M.J., Newton R.U., Triplett-McBride T. et al. (2001). Changes in myosin heavy chain composition with heavy resistance training in 60- to 75-year-old men and women. Eur. J. Appl. Physiol.84 (1-2):127-132.
132. Shen W., Zhang X., Zhao G. et al. (1995). Nitric oxide production and NO synthase gene expression contribute to vascular regulation during exercise. Med. Sci. Sports Exerc.27(8):1125-1134.
133. Shimano T., Kraemer W.J., Spiering B.A., et al. (2006). Relationship between the number of repetitions and selected percentages of one repetition maximum in free weight exercises in trained and untrained men. J. Strength Cond. Res. 20 (4):819-823.
134. Simoes R.P., Mendes R.G., Castello V., et al. (2010). Heart-rate variability and blood-lactate threshold interaction during progressive resistance exercise in healthy older men. J. Strength Cond. Res.24(5):1313-1320.
135. Smolle M., Prior A.E., Brown A.E. et al. (2006). A new level of architectural complexity in the human pyruvate dehydrogenase complex. J. Biol. Chem.281 (28): 19772-19780.
136. Spriet L.L., Soderiund K., Bergstrom M., et al. (1987). Anaerobic energy release in skeletal muscle during electrical stimulation in men. J. Appl. Physiol.62 (2):611-615.
137. Staron R.S., Hikida R.S., Hagerman F.C. et al. (1984). Human skeletal muscle fiber type adaptability to various workloads. J. Histochem. Cytochem.32(2): 146-152.
138. Staron R.S., Karapondo D.L., Kraemer W.J. et al. (1994). Skeletal muscle adaptations during early phase of heavy resistance traiing in men and women. J. Appl. Physiol.76(3): 1247-1255.
139. Steding K., Engblom H., Buhre T., Carlsson M. et al. (2010). Relation between cardiac dimensions and peak oxygen uptake. J. Cardlovasc. Magn. Reson. 12 (8): 1-9.
140. Stewart J.M., Medow M.S., Montgomery L.D. et al. (2004). Decreased skeletal muscle pump activity in patients with postural tachycardia syndrome and low peripheral blood flow. Am. J. Physiol. Heart Circ. Physiol.286(3):H1216-1222.
141. Storen 0., Helgerud J., Stoa E.M. et al. (2008). Maximal strength training improves running economy in distance runners. Med. Sci. Sports Exerc.40:1089-1094.
142. Stromme S.B., Ingjer F., Meen H.D. (1977). Assessment of maximal aerobic power in specifically trained athletes. J. Appl. Physiol.42(6): 833-837.
143. Sweet T.W., Foster C., McGuigan M.R., et al. (2004). Quantitation of Resistance Training using the session rating of perceived exertion method. J. Strength Cond. Res.18 (4):796-802.
144. Tanaka H., Swensen T. (1998). Impact of resistance training on endurance performance: A new form of cross-training? Sports Med. 25(3):191-200.
145. Tang J.E., Hartman J.W., Phillips S.M. (2006). Increased muscle oxidative potential following resistance training induced fiber hypertrophy in young men. Appl. Physiol. Nutr. Metab. 31 (5) 495-501.
146. Tanimoto M., Kawano H., Gando Y. et al. (2009). Low-intensity resistance training with slow movement and tonic force generation increase basal limb blood flow. Clin. Physiol. Funct. Imaging.29:128-135.
147. Tarnopolsky M.A. (2009). Mitochondrial DNA shifting in older adults following resistance exercise training. Appl. Physiol. Nutr. Metab. 34 (3):348-354.
148. Tesch P.A., Colliander E.B., Kaiser P. (1986). Muscle metabolism during intense, heavy resistance exercise. Eur. J. Appl. Physiol. Occup. Physiol.555 (4):362-366.
149. Tesch P.A., Thorsson A., Essen-Gustavsson B. (1989). Enzyme activities of FT and ST muscle fibers in heavy-resistance trained athletes. J. Appl. Physiol.67(1):83-87.
150. Tinken T.M., Thijssen D.H., Hopkins N., et al. (2010). Shear stress mediates endothelial adaptations to exercise training in humans. Hypertension.55(2):312-318.
151. Urhausen A., and Kindermann W. (1999). Sports-specific adaptations and differentiation of the athletes' heart. Sports Med. 28 (4):237-244.
152. Walker P.M., Brunotte F., Rouhier-Marcer I. et al. (1998). Nuclear magnetic resonance evidence of different muscular adaptations after resistance training. Arch. Phys. Med. Rehabil.79 (11): 1391-1398.
153. Weber M.A., Hildebrandt W., Schroder L. et al. (2010). Concentric resistance training increases muscle strength without affecting microcirculation. Eur. J. Radiol.73(3):614-621.
154. Wilmore J.H., Costill D.L., Kenney W.L. (2008). Physiology of Sport & Exercise (4th Edition). Champaign: Human Kinetics.
155. Willoughby D.S., Pelsue S. (2000). Effects of high intensity strength on steady state myosin heavy chain isoform mrna expression. JEPonline.3 (4): 13-25.
156. Wray D.W., Donate A.J., Nishiyama S.K. et al. (2007). Acute sympathetic vasoconstriction at rest and during dynamic exercise in cyclists and sedentary humans. J. Appl. Physiol.102 (2):704-712.
157. Wu Z., Puigserver P., Andersson U. et al. (1999). Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell.98 (1): 115-124.
Cita Original
Steele J, Fisher J, McGuff D, Bruce-Low S, Smith D. (2012) .Resistance Training to Momentary Muscular Failure Improves Cardiovascular Fitness in Humans: A Review of Acute Physiological Responses and Chronic Physiological Adaptations. JEPonline 15(3):53-80.
Cita en PubliCE
James Steele, James Fisher, Doug McGuff, Stewart Bruce-Low Dave Smith (2015). El Entrenamiento de la Fuerza Hasta el Fallo Mejora la Aptitud Cardiovascular en Humanos: Revisión de las Respuestas Fisiológicas Agudas y Adaptaciones Fisiológicas Crónicas. .https://g-se.com/el-entrenamiento-de-la-fuerza-hasta-el-fallo-mejora-la-aptitud-cardiovascular-en-humanos-revision-de-las-respuestas-fisiologicas-agudas-y-adaptaciones-fisiologicas-cronicas-1818-sa-257cfb272537fd