Determinantes de la Variabilidad de la Tasa de Intercambio Respiratorio en Reposo y Durante el Ejercicio en Atletas Entrenados
Timothy D Noakes, Julia H Goedecke, Liesl Grobler, Malcolm Collins, Estelle V Lambert, Alan St Clair Gibson0 University of Cape Town Bioenergetics of Exercise Research Unit, University of Cape Town Medical School, Newlands 7725, Sudáfrica.
Resumen
Analizamos la variabilidad y los determinantes de la tasa de intercambio respiratorio (RER) en reposo y durante el ejercicio en 61 ciclistas entrenados. Se midió la RER en ayuno (10-12 h) en reposo y durante el ejercicio a 25, 50 y 70% de la producción de potencia máxima (Wmax), y además se tomaron muestras de sangre para determinar la concentración de lactato y de ácidos graso libres (FFA). Antes de realizar estas mediciones, se determinó el volumen de entrenamiento, la ingesta dietética, la composición de fibras musculares, la concentración de sustrato y las actividades enzimáticas. Se observó una gran variabilidad entre los individuos en la RER en reposo (0,718-0,927) que persistió durante los ejercicios de intensidad creciente. Los principales determinantes de RER en reposo, incluyeron el contenido de glucógeno en el músculo, el volumen de entrenamiento, proporción de fibras Tipo 1, [FFA] y [lactato] y % de ingesta de grasas en la dieta (r2 ajustado= 0,59, P <0,001). A excepción de la composición de fibras musculares, estas variables también sirvieron para la estimación de la RER a 25, 50 y 70% de Wmax en diferentes grados. El determinante clave al 25% de Wmax fue la concentración de sustratos provenientes de la sangre, al 50% fue la concentración de sustrato en el músculo y la actividad de las enzimas glucolíticas y al 70% fue la concentración de lactato. La RER de reposo también fue un determinante significativo de RER al 25 % (r = 0,60) y 50% (r = 0,44) de Wmax.
Palabras Clave: actividad enzimática muscular, contenido de glucógeno, triacilglicéridos, volumen de entrenamiento, ingesta dietaria
INTRODUCCION
Hace mucho tiempo se sabe que, tanto la grasa como los carbohidratos (CHO) de la dieta sirven como sustratos para el metabolismo energético en sujetos que realizan ejercicios y que la contribución relativa de estos dos sustratos para impulsar la producción de potencia puede ser influenciada por factores como la dieta previa al ejercicio (3, 5, 38, 39), el nivel de entrenamiento (3, 6, 8, 28), intensidad del ejercicio (3, 6, 37), y el entorno hormonal relativo durante el ejercicio (17).
Sin embargo, también existiría variabilidad individual en la oxidación de los sustratos durante el ejercicio, tanto en sujetos desentrenados (22, 43) como en sujetos entrenados (5, 36). Esta variabilidad en la utilización de los sustratos durante el ejercicio puede estar relacionada con las diferencias en las características del músculo esquelético. De hecho, Wade et al. (43) observaron que la tasa de intercambio respiratorio (RER) durante el ejercicio moderado (100 W) en sujetos alimentados, estaba inversamente asociada con la proporción de fibras musculares de tipo I (contracción lenta). Recientemente, Geerling et al. (18) y Helge et al. (22) no lograron corroborar estos resultados en sujetos desentrenados que realizaron ejercicios con la misma intensidad relativa [55% de intensidad relativa de ejercicio (VO2máx)] en condición de ayuno. Además, Zurlo et al. (47) tampoco lograron demostrar una relación entre la RER de 24-h (24hRER) y la composición de fibras musculares en una muestra pequeña de hombres y mujeres sedentarios no obesos. Sin embargo, ellos observaron una correlación inversa entre la 24hRER y la actividad de la β-hidroxiacil-CoA deshidrogenasa en el músculo, incluso después de realizar ajustes para la edad, sexo y % de grasa corporal [r = -0,70, P = 0,005 (47)].
Los trabajos adicionales que estudiaron la variabilidad y determinantes de utilización de sustratos se realizaron en sujetos desentrenados, a menudo obesos, alimentados con una dieta estandarizada en una cámara metabólica. En un estudio reciente realizado con 71 hermanos saludables de 31 familias, Toubro et al. (42) observaron que una parte sustancial de la variación entre los individuos en la 24hRER podía ser explicada por la edad, género, balance energético, ingesta dietética actual, y concentraciones de insulina y ácidos grasos libres (FFA) en el plasma. Sin embargo, ellos también encontraron que había una similitud familiar fuerte en 24hRER, lo que había sido previamente demostrado por Zurlo et al. (46).
Sin embargo, no conocemos ningún trabajo, que haya estudiado en forma exhaustiva la variabilidad y los posibles determinantes de la utilización de sustratos tanto en reposo como durante el ejercicio en atletas entrenados que consumen sus dietas habituales. La variabilidad en la utilización de sustratos en los atletas entrenados sólo ha sido analizada de manera indirecta tal como se ha demostrado en dos estudios recientes de este laboratorio (5, 36). Estos estudios demostraron que no todos los ciclistas entrenados podían aumentar la contribución de las grasas al metabolismo oxidativo durante pruebas prolongadas de ejercicio asociados con el agotamiento de glucógeno muscular (5, 36). Los ciclistas que no aumentaban sus tasas relativas de oxidación de grasas durante ejercicios prolongados en estado estable tenían mayores tasas iniciales y globales de oxidación de CHO y alcanzaban la fatiga antes que los ciclistas que tenían mayores tasas relativas de oxidación de grasas (y menores tasas de oxidación de CHO)
Por consiguiente, el objetivo de este trabajo fue, en primer lugar, estudiar la variabilidad en la utilización de sustratos (RER) del cuerpo entero en ayuno, en reposo y durante la realización de ejercicios en estado estable en diferentes intensidades, en un grupo de ciclistas con capacidad de rendimiento superior a la del promedio. Y en segundo lugar, este estudio intentó identificar las relaciones entre la utilización de sustratos durante el reposo y durante el ejercicio, así como también los posibles determinantes de RER, entre los que se incluyen las características del músculo esquelético, entrenamiento, ingesta dietaria y composición corporal.
METODO
Selección de Sujetos y Muestreos
Cuarenta y cinco varones y dieciséis mujeres, con entrenamiento en ciclismo de resistencia participaron en esta investigación, que fue aprobada por el Comité de Ética e Investigación de la Facultad de Ciencias de Salud de la Universidad de Ciudad del Cabo. Para participar en el estudio, los sujetos debían: 1) haber completado una carrera de ciclismo local de 104-km en <3,5horas en el caso de los varones y en <4 h en el caso de las mujeres y 2) no tener ninguna enfermedad metabólica conocida que pudiera haber afectado adversamente el metabolismo intermediario (por ejemplo, diabetes, desórdenes de las hormonas tiroideas, hiperlipidemia). Los sujetos no estaban consumiendo ninguna medicación para condiciones crónicas como tensión arterial alta (por ejemplo, (antagonistas de los receptores β-adrenérgicos) ni estimulantes para afecciones como el asma (por ejemplo, agonistas de los receptores β-adrenérgicos). Los participantes fueron informados de la naturaleza de la evaluación y se les explicaron todos los riesgos y beneficios potenciales. Antes de comenzar el estudio los participantes dieron su consentimiento informado por escrito.
En el estudio se incorporaron mujeres para caracterizar cualquier diferencia, entre varones y mujeres, en la utilización de sustratos en reposo y durante el ejercicio. Debido a inconvenientes de tiempo y logística no pudimos realizar control sobre el ciclo menstrual en las mujeres.
Evaluaciones Preliminares
Antropometría
Se realizaron las mediciones antropométricas, entre las que se incluyeron la suma de siete pliegues cutáneos (bíceps, tríceps, sub-escapular, suprailíaco, abdomen, muslo y pantorrilla), perímetro de cintura y cadera y masa muscular. El porcentaje de grasa corporal se estimó por medio de las ecuaciones de Durnin y Womersley (13).
Producción de Potencia Máxima y Consumo de Oxígeno Máximo
El consumo de oxígeno máximo (VO2máx) y la producción de potencia máxima sostenida (Wmax) se determinaron en una bicicleta ergométrica con freno electrónico (Lode, Groningen, Holanda) modificada con punteras y manillar de carrera, tal como lo describieron previamente Hawley y Noakes (21). La tasa de trabajo comenzó en 3,33 W/kg de masa corporal para los varones y 150 W para las mujeres. Después de 150 s, el trabajo se incrementó 50 W y luego 25 W cada 150 s hasta que los sujetos se sintieran exhaustos. El agotamiento fue definido como una reducción >10% en la frecuencia de pedaleo, o RER >1,10, o ambos. Wmax se definió como la mayor intensidad de ejercicio que los sujetos completaron en 150 s, en W, más la fracción de tiempo transcurrido en la tasa de trabajo final multiplicada por 25 W. Los valores de Wmax fueron usados para determinar las cargas de trabajo relativas en las pruebas experimentales subsiguientes.
Durante la prueba de esfuerzo progresiva, se midieron el volumen de ventilación, consumo de oxígeno (VO2), y producción de CO2 (VCO2) en intervalos de 15s mediante un Analizador respiración por respiración, Oxycon Alfa (Jaeger, Wuerzburg, Netherlands). Antes de cada prueba, el medidor de gases fue calibrado con una jeringa de 3 litros Hans Rudolph (Vacumed, Ventura, CA), y los analizadores fueron calibrados con el aire del cuarto y una mezcla de gases de 4% CO2-96% N2.
Historial de Entrenamiento
Cada sujeto proporcionó un historial de entrenamiento retrospectivo detallado. Además, los participantes completaron un registro de entrenamiento durante las 2 semanas previas a la prueba experimental. El entrenamiento fue cuantificado en equivalentes metabólicos (METs, por semana) según la intensidad y duración.
Análisis de la Dieta
Tres días antes de la prueba experimental, los participantes completaron un registro dietario con alimentos pesados. Los archivos dietéticos se analizaron con el programa Food Finder (Medtech, Medical Research Council, Tygerberg, South Africa) para determinar la ingesta de energía y el consumo de macronutrientes de los sujetos.
Prueba Experimental
Tipo de Fibra Muscular, Contenido de Sustrato y Actividades Enzimáticas
El día antes de la prueba experimental, se instruyó a los sujetos para que entrenaran de manera habitual durante no más de 1 hora antes de las 18:00 hs. En el día posterior, los sujetos acudieron al laboratorio después de un ayuno de 10 a 12-h durante la noche. Se realizaron biopsias musculares en reposo del músculo vasto lateral (vastus lateralis) de 56 sujetos mediante la técnica de biopsia percutánea con aguja. Una porción de la muestra de músculo fue rápidamente congelada en N2 líquido y almacenada a -80°C para el posterior análisis de contenido de glucógeno y triacilglicéridos, y para la determinación de las actividades enzimáticas entre las que se incluían las actividades de carnitina aciltransferasa (CAT), citrato sintetasa (CS), fosfofructoquinasa (PFK), hexoquinasa (HK), β-hidroxiacil-CoA deshidrogenasa (β-HAD). La muestra restante fue orientada y embebida en Tissue-Tek (Miles Laboratories, Naperville, IL), congelada en n-pentano enfriado en nitrógeno líquido (Saarchem, Muldersdrift, Sudáfrica), y fue almacenado -20°C para la posterior determinación del tipo de fibra mediante el método de la ATPasa miofibrilar (mATPasa). Brevemente, se obtuvieron secciones seriales (10 µm para el pH 9,4 y 20 µm para el pH 4,3 y 4,6) mediante un criostato a -20°C. Las secciones adyacentes de músculo fueron analizadas por ATPasa miofibrilar a pH 9,4 después de una preincubación acida (el pH 4,3 o 4,6) y alcalina (pH 10,4) para identificar los tres tipos principales de fibras (12).
Antes de efectuar el análisis bioquímico del contenido de glucógeno y triacilglicéridos del músculo, una porción de la muestra de biopsia de músculo congelada (~50 mg) fue deshidratada por congelación y fue disecada libre de cualquier rastro de grasa o de tejido conjuntivo visible. El contenido de glucógeno muscular se determinó en función de los residuos de glucosa (método de glucosa oxidasa; Analizador de Glucosa 2, Beckman Instruments, Fullerton, CA) luego de la hidrólisis de la muestra de músculo en HC1 2M a 95°C durante 3 h (35). Las concentraciones de glicerol se determinaron con un equipo comercial de determinación de glicerol (Boehringer Mannheim, Mannheim, Alemania) después de que los triglicéridos fueran degradados a glicerol y ácidos grasos libres (FFAs), tal como lo describieran Kiens y Richter (30).
Para la determinación de las actividades enzimáticas en el músculo, 15-50 mg (peso húmedo) de músculo fueron homogeneizados en buffer fosfato en hielo (1:19 peso/vol) y sonicados en hielo 3 X 10 s (Virsonic 60, El Vitrus, Nueva York, NY). Se determinaron las actividades de CAT y PFK utilizando el método de Crabtree y Newsholme (10) y Ling et al. (32), respectivamente. HK y β-HAD fueron determinadas mediante las técnicas descritas por Bass et al. (2). La actividad de la CS se determinó mediante la técnica descrita por Srere (40). Las actividades enzimáticas se expresaron en relación al contenido de proteínas y fueron determinadas por duplicado a través del Ensayo de determinación de proteínas de Bio-Rad (Laboratorios Bio-Rad, Munich, Alemania), utilizando albúmina de suero bovino como estándar.
RER en Estado Estable
Después de que se obtuvieran las biopsias del músculo, los sujetos descansaron durante 30 min y/o hasta que su frecuencia cardíaca retornara a niveles previos a la biopsia. Luego se determinaron los valores de VO2, VCO2 y RER durante 15 min en reposo y durante ejercicios de ciclismo en estado estable a 25, 50 y 70% de Wmax, respectivamente en los cuales se mantuvo una frecuencia de pedaleo de 90 rpm. Estas cargas de trabajo correspondían a 41, 63 y 80% de VO2max, respectivamente. Las mediciones de intercambio de gases fueron registradas durante 5 min en reposo y en cada carga de trabajo del ejercicio, después de un "período de estabilización" de 10-min, tal como se describiera anteriormente.
La confiabilidad del Analizador Oxycon Alfa fue evaluada semanalmente quemando etanol absoluto [reactivo analítico con 99% de pureza, Associated Chemical Enterprises (Pty), Glenvista, Sudáfrica] como referencia. La confiabilidad de RER, en reposo y durante el ejercicio en las diferentes intensidades del ejercicio, fue evaluada previamente en nuestro laboratorio (observaciones no publicadas). El coeficiente de variación (CV) para RER en reposo, evaluado en 13 varones saludables en tres ocasiones, fue 2,1%. Los CV para RER a 25, 50 y 70% de Wmax, evaluado en nueve ciclistas varones entrenados en resistencia en tres ocasiones, fue 2,0, 1,5 y 1,4%, respectivamente. Los coeficientes de correlación intraclase para RER en reposo y en 25, 50 y 70% Wmax fueron 0,847, 0,658, 0,814 y 0,843, respectivamente.
Muestreo y Análisis de la Sangre
Las muestras de sangre (~6 ml) fueron obtenidas de una vena del antebrazo en reposo y durante el último minuto de cada carga de trabajo. Una alícuota (3 mL) fue colocada en un tubo que contenía oxalato de potasio y fluoruro de sodio para el análisis subsecuente de concentraciones plasmáticas de lactato. La alícuota restante (3 ml) se colocó en un tubo que contenía gel y activador de coagulación para las determinaciones las concentraciones séricas de ácidos grasos libres (FFA, no esterificados). Todas las muestras fueron mantenidas en hielo hasta el momento en que fueron centrifugadas a 3000 rpm a 4°C durante 10 min al finalizar la prueba. El plasma y el suero fueron luego almacenados a -20°C para los análisis posteriores.
Las concentraciones plasmáticas de lactato (Lactate Pap, Bio Merieux, Marcy-L Etiole, France) y las concentraciones séricas de FFA (FFA Half-micro test, Boehringer Mannheim) fueron determinadas mediante mediciones espectrofotométricas enzimáticas con equipos con reactivos comerciales.
Análisis Estadísticos
Se usaron correlaciones bivariadas para analizar las relaciones entre el RER en estado estable, en reposo y durante el ejercicio, y las características fisiológicas, características musculares, entrenamiento y dieta. Luego estas variables fueron utilizadas para el análisis multivariado, por medio de la técnica de eliminación progresiva, para determinar el modelo que mejor predice la RER en reposo y durante el ejercicio en las diferentes intensidades. Un ANOVA de mediciones repetidas se utilizó para investigar las diferencias en RER entre varones y mujeres. Todos los resultados se presentan como Media±DS, y el nivel de significancia estadística se fijó en un nivel de α de p<0,05 .
RESULTADOS
Características de los Sujetos
Las características sujetas se presentan en la Tabla 1. Aunque los sujetos realizaron ejercicio en niveles similares (completaron una carrera de ciclismo de 104-km en el 20% superior del campo), se observó una gran variación entre los sujetos en la composición corporal y la capacidad de trabajo físico (Tabla 1). En la Tabla 1 también se presentan los informes de los sujetos sobre ingesta energética y de macronutrientes durante los 3 días previos a la prueba experimental. Dado que no se observaron diferencias entre los varones y mujeres en el valor de RER en reposo y durante el ejercicio a 25, 50 y 70% de Wmax (Figura 1), todos los datos se combinaron para todos los análisis realizados en este estudio.
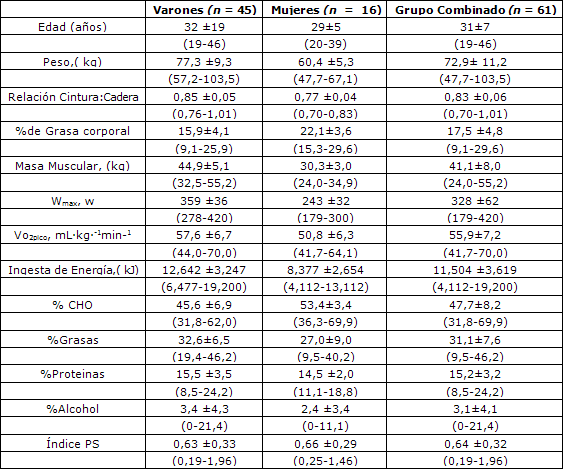
Tabla 1. Características de los sujetos. Los valores de expresan en forma de Media± SD; con los intervalos de valores entre paréntesis. Wmax= producción de potencia máxima; VO2max= Consumo de oxígeno máximo; CHO= carbohidratos; Indice PS= Relación entre grasas poli-insaturadas y saturadas.
Figura 1. Tasa de intercambio respiratorio (RER; Media± SD) en reposo y durante ejercicios realizados en diferentes intensidades en varones (n=45) y mujeres (n=16). VO2 = Consumo de oxígeno; Wmax = producción de potencia máxima.

Variabilidad en RER
La RER en reposo alcanzó valores de 0,927 a 0,718 que corresponden a una tasa relativa de oxidación de grasas de 23 a 93% (Fig. 2A). A medida que la intensidad del ejercicio aumentaba, se observó un aumento correspondiente en la RER media (Figura 2B).
La RER media se incrementó de 0,817 ± 0,051 en reposo a 0,862 ± 0,037, 0,900 ± 0,040, y 0,976 ± 0,043 en 25, 50, y 70% Wmax, respectivamente. Sin embargo, la variabilidad en RER se mantuvo durante el ejercicio en todas las intensidades, y con RER tomando valores comprendidos entre 0,818 y 0,983 (Fig. 2B) durante el ejercicio sub-máximo (50% Wmax).
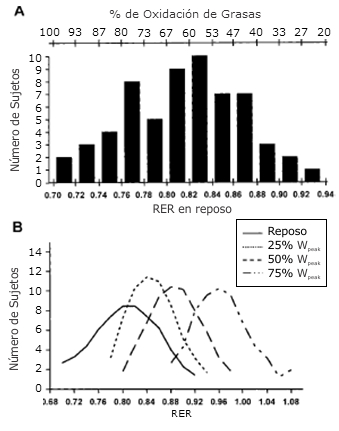
Figura. 2. Distribución de frecuencias de la RER en ayuno y reposo (A) y durante la realización de ejercicio en estado estable al 25, 50 y 70% de Wmax (B).
Correlaciones con RER en Reposo y Durante el Ejercicio
Para describir la relación entre RER en reposo y durante el ejercicio y los diferentes parámetros fisiológicos y metabólicos se aplicó un análisis bivariado (Tabla 2). Las variables que se correlacionaron de manera positiva con la RER en reposo fueron la proporción de fibras musculares de Tipo I, el contenido de glucógeno en el músculo en reposo y las concentraciones plasmáticas de lactato. La proporción de fibras musculares de Tipo IIa, concentraciones de FFA en el suero y la ingesta dietaria de grasas se correlacionó de manera negativa con la RER en reposo. La RER en reposo también se correlacionó significativamente con RER tanto al 25 como al 50% Wmax (r = 0,60, P <0,0001, y r = 0,44, P <0,0001, respectivamente) pero no al 70% Wmax (r = 0,17, no significativo).
Al 25% de Wmax, la composición de fibra muscular y el contenido de glucógeno muscular no se correlacionaron con RER. Sin embargo, las concentraciones séricas de FFA y de lactato en el plasma, la ingesta dietaria de grasas y el volumen de entrenamiento se correlacionaron significativamente con RER. Al 50% de Wmax, las concentraciones séricas de FFA y de lactato en el plasma, la ingesta dietaria de grasas y el volumen de entrenamiento se correlacionaron significativamente con RER (Tabla 2). Por otra parte, el contenido muscular de triacilglicéridos y la actividad de CS se correlacionaron de manera negativa con la RER durante el ejercicio, mientras que las proporciones PFK-a-CS y HK-a-CS (PFK/CS y HK/CS), que representan ambas el flujo glucolítico, se correlacionaron positivamente con la RER al 50% de Wmax. Al 70% Wmax, sólo se observó una asociación significativa con RER y las concentraciones plasmáticas de lactato, HK/CS y PFK/CS y el volumen de entrenamiento (Tabla 2).
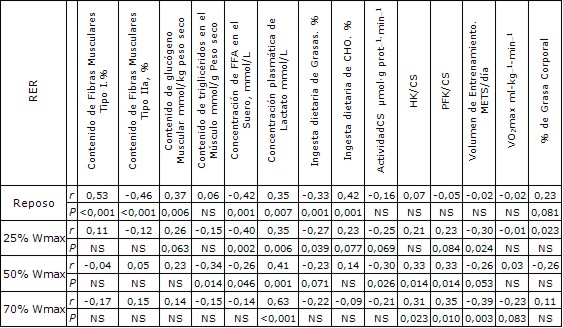
Tabla 2. Matriz de correlación para la RER en ayuno en condición de reposo y durante los ejercicios en estado estable en diferentes intensidades.
Análisis de Multivariado para la Determinación de RER en reposo
Las variables, proporción de fibras musculares tipo I, contenido de glucógeno muscular, volumen de entrenamiento, ingesta dietaria de grasas y concentraciones séricas de FFA en reposo y concentración de lactato en el plasma fueron incluidas en el modelo que explicó el 59% de la varianza en la RER en reposo [P <0,0001, error estándar de la estimación (SEE) = 0,035, Tabla 3]. La proporción de fibras musculares de tipo I se asoció de manera positiva con la RER en reposo y fue el determinante más importante de RER en esta ecuación.
Además de la composición de fibras de músculo, el contenido de glucógeno muscular también se comportó como determinante importante de la RER en reposo en esta ecuación. El volumen de entrenamiento, la concentración de FFA en reposo, y la ingesta dietaria de grasas, presentaron una correlación menor, pero aún así se comportaron como determinantes significativos de la RER en reposo. Las actividades enzimáticas musculares, el contenido de triacilglicéridos del músculo, el porcentaje de grasa corporal, y VO2max no fueron estimadores significativos de RER en reposo y por lo tanto no fueron incluidos en el modelo de predicción de RER en reposo.
Determinantes de RER durante Ejercicios de Baja a Moderada Intensidad
El modelo estadístico que el mejor predijo la RER al 25% de Wmax todavía incluía al glucógeno muscular y el volumen de entrenamiento como determinantes importantes, pero no incluía el tipo de fibra muscular. Por otra parte, la actividad muscular de CS, las concentraciones séricas de FFA y la ingesta dietética de grasas contribuyeron significativamente con el modelo. Cuando estas variables fueron incorporadas en la ecuación, explicaron el 45% de la varianza en RER al 25% de Wmax (P <0,0001, SEE= 0,029, Tabla 4). Sin embargo, cuando la RER en reposo se utilizó en la ecuación como una variable, el R2 ajustado aumentó a 0,575 (P <0,0001, SEE = 0,029, Tabla 4). En este modelo, el volumen de entrenamiento y el contenido de glucógeno muscular se volvieron los determinantes menos importantes de RER al 25% al Wmax, pero aumentó la importancia relativa de las enzimas del músculo, entre las que se incluían las actividades de CS y de PFK.
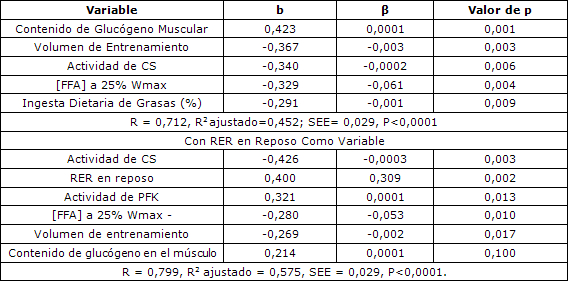
Tabla 3. Análisis multivariado para la RER en reposo. R = 0,797, R2 ajustado = 0,585, SEE = 0,035, P <0,0001. β= estimador del parámetro; b= coeficiente de la correlación parcial.
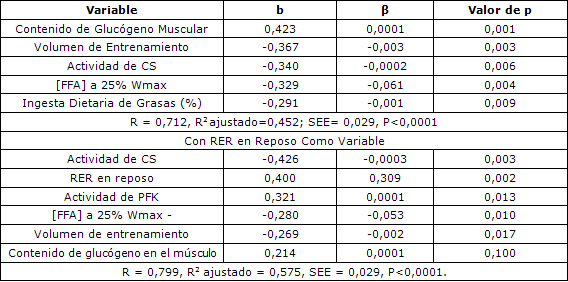
Tabla 4. Análisis multivariado para RER al 25% de Wmax.
Aunque la intensidad de la serie de ejercicios se duplicó de 25 al 50% de Wmax, se observó que las variables que mejor predecían la RER eran similares (Tabla 5). Sin embargo, en esta mayor intensidad de ejercicio, el nivel de triacilglicéridos en el músculo se volvió un estimador significativo de RER, mientras que las concentraciones de FFA circulantes contribuyeron menos al modelo que en el caso del 25% de Wmax. Este modelo explicó el 42% de la varianza de RER al 50% de Wmax (P <0,0001, SEE = 0,031, Tabla 5).
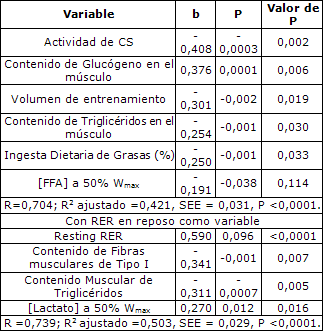
Tabla 5. Análisis de multivariado para RER al 50% de Wmax. [Lactato]= Concentración de Lactato; [FFA]= Concentración de Ácidos Grasos Libres.
Cuando la RER en reposo se incorporó como variable en el modelo para predecir RER al 50% de Wmax, el ajuste del modelo mejoró (r2 ajustado = 0,50, SEE = 0,029, P <0,0001; Tabla 5). Otras variables en la ecuación incluyeron la proporción de fibras musculares de tipo I, contenido de triacilglicéridos en el músculo, y variables menos importantes como las concentraciones plasmáticas de lactato. Notablemente, la proporción de fibras musculares de tipo I fue un mejor factor de predicción de la RER al 50% que las fibras musculares de tipo IIa (b ponderado = -0,47 y -0,16 para proporción de fibras de tipo I y tipo IIa, respectivamente).
Determinantes de RER Durante Ejercicios de Alta-intensidad
Durante el ejercicio de alta-intensidad (70% Wmax), la concentración plasmática circulante de lactato contribuyó significativamente con el modelo de predicción de la RER (Tabla 6). Este modelo que también incluyó variables como la ingesta dietaria de grasas, volumen de entrenamiento, contenido de glucógeno en el músculo y concentración circulante de FFA, explicó el 56% de la varianza RER (P <0,0001, SEE = 0,031, Tabla 6). La ingesta de grasas en la dieta y el volumen de entrenamiento, presentaron una asociación negativa con RER y contribuyeron de manera similar con el modelo, mientras que el contenido de glucógeno en el músculo se asoció positivamente con RER pero su contribución en el modelo fue menor.
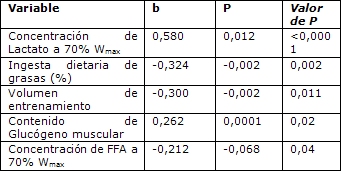
Tabla 6. Análisis multivariado de RER al 70% de Wmax.
Posibles Variables de Confusión
Hay diferentes variables que presentaron asociación entre ellas y podrían haber confundido la interpretación de las relaciones con RER descriptas anteriormente. La edad se correlacionó significativamente con el porcentaje de grasa corporal y con el VO2max (r = 0,34, P = 0,007 y r = -0,31, P = 0,015, respectivamente). El porcentaje de grasa corporal se correlacionó negativamente con el VO2max (r = -0,75, P <0,001) y positivamente con el contenido muscular de triacilglicéridos (r = 0,30, P = 0,03). El volumen de entrenamiento se correlacionó de manera positiva con la actividad de CS y el contenido de glucógeno en el músculo (r = 0,30, P = 0,028 y r = 0,36, P = 0,011, respectivamente) y presentó una tendencia a correlacionarse con la proporción de fibras musculares de tipo I (r = 0,26, P = 0,055). En la Tabla 7 se presentan las correlaciones relacionadas a las características del músculo.
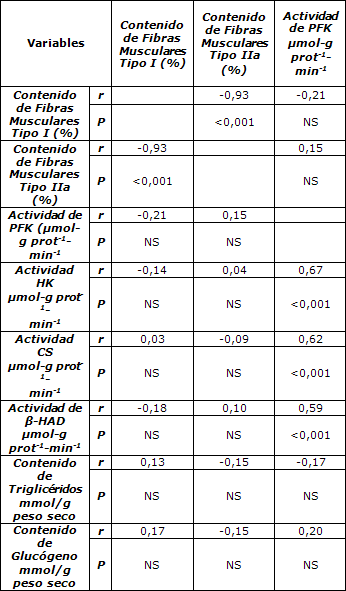
Tabla 7. Matriz de correlación de las variables relacionadas con el músculo. β-HAD= β-hidroxiacil Co-A deshidrogenasa.
DISCUSION
Variabilidad de RER en Ayuno, en Reposo y Durante los Ejercicios en Estado Estable a Corto Plazo
El primer hallazgo importante de este estudio fue la gran variabilidad de RER en reposo y durante el ejercicio en las diferentes intensidades en los atletas entrenados en resistencia. La tasa relativa de oxidación de grasas en reposo tomó valores de 23 a 93%, que es casi una diferencia de cuatro veces en la oxidación de grasas en estos atletas con capacidad de rendimiento por encima del promedio (Fig. 2A).En trabajos previos se ha demostró una variabilidad similar en la RER en reposo o en la 24hRER (0,771-0,931) en sujetos desentrenados (33, 45-47). Recientemente, Helge et al. (22) informaron una gran variabilidad en RER (0,83-0,95) durante el ejercicio en estado estable de baja intensidad. Sin embargo, según nuestros conocimientos, éste es el primer estudio en el cual se describió la gran variabilidad en la utilización de sustratos en reposo y durante el ejercicio, en atletas entrenados en resistencia que poseían una capacidad de rendimiento superior a la del promedio. Por otra parte, nosotros no conocemos ningún otro estudio que haya demostrado que esta variabilidad en la RER en reposo persiste en ejercicios de baja, moderada y alta-intensidad de corto plazo.
Un segundo hallazgo importante de este estudio, que no ha sido previamente reportado, fue que la distribución normal de RER en reposo se mantuvo durante los ejercicios de intensidad creciente (Figura 2B). La distribución normal se observó hasta en la intensidad de ejercicio relativamente alta de 70% Wmax.
Sin embargo, nosotros no podemos descartar la posible influencia del propio procedimiento de biopsia muscular en las mediciones respiratorias y oxidación de sustratos. Hay pocos datos que sugieren que las concentraciones de hormonas de estrés como el cortisol y la norepinefrina pueden permanecer elevadas en el plasma durante ≥20 min después de la biopsia (22, 24). Sin embargo, en estos estudios, los cambios hormonales no se asociaron directamente con los cambios en las concentraciones de metabolitos plasmáticos en ayuno (22, 24). Además, nuestras mediciones fueron realizadas luego de cómo mínimo 40 min después de la biopsia y fueron realizadas en todo el grupo, minimizando así el posible sesgo.
Determinantes de la RER en Ayuno en Reposo
Los determinantes de RER en reposo incluyeron la proporción de fibras musculares de tipo I, contenido de glucógeno muscular, la ingesta dietaria de grasas, entrenamiento y los metabolitos en la sangre (lactato plasmático y concentraciones de FFA en el suero), y entre todos explicaron aproximadamente el 56% de la varianza de RER (Tabla 3). No esperábamos la asociación positiva con las fibras musculares de tipo I (contracción lenta) y la asociación negativa con las fibras musculares de tipo Ila (rápidas-oxidativas) y la RER. Las fibras musculares de tipo I, a menudo utilizan más los ácidos grasos como fuente de combustible, dado que tienen una elevada capacidad oxidativa gracias a una elevada densidad mitocondrial y capilar (26). Mas aún, Wade et al.. (43) observaron que la RER, medida durante el ejercicio moderado a una intensidad absoluta de 100 W en 11 varones sedentarios, se relacionó inversamente con la proporción de fibras musculares de tipo I. Sin embargo, Zurlo et al. (47), y más recientemente, Helge et al. (22) no observaron ninguna asociación entre la composición de fibras musculares y RER en sujetos desentrenados.
Nosotros no podemos explicar totalmente las razones para la asociación positiva entre la proporción de fibras musculares de tipo I y la RER en reposo en este estudio. Una posible explicación podría ser la coexistencia de isoformas de la cadena pesada de miosina para las fibras de tipo I y Ila encontradas en los sujetos entrenados en resistencia pero no en los individuos sedentarios (31). Si existe una gran coexistencia de isoformas de tipo I y tipo II en las muestras de músculo, tal como se esperaba en estos ciclistas entrenados, la técnica convencional de tipificación de fibras mediante la ATPasam podría haber arrojado una reacción histoquímica como la de las fibras de tipo I, sobrestimando la proporción de fibras de tipo I. Sin embargo, nosotros no podemos excluir la posibilidad que la relación entre el tipo de fibra muscular y la RER, pueda haber sido influenciada por otro/s factor/es no identificados en este estudio.
Asociación Entre RER y Composición Corporal
Nosotros no encontramos ninguna asociación entre la RER en ayunas y en reposo y la composición corporal tal como lo demostraron Zurlo et al. (46) y Weyer et al. (45) en sujetos desentrenados, que realizaron las determinaciones mediante técnicas de pesaje hidrostático y absorciometría de rayos X de energía dual para todo el cuerpo, respectivamente (Tabla 2). La falta de asociación entre RER y la composición corporal puede deberse al hecho que nuestros sujetos eran mas magros y tenías un menor intervalo de adiposidad corporal (9-30%) en comparación con los sujetos de los estudios de Zurlo et al. (46) (5-50%) y Weyer et al. (45) (3-53%). Tampoco observamos ninguna asociación entre RER durante el ejercicio y la composición corporal, previamente descripta por Wade et al. (43) (r = 0,538, P <0,001) en 50 varones con un intervalo de adiposidad corporal similar al de los sujetos de nuestro estudio (~4-28%). Sin embargo, sus participantes no estaban entrenados y no se encontraban en ayunas cuando se realizó la medición de RER durante un ejercicio moderado con la misma intensidad relativa pero no absoluta de ejercicio (100 W). En concordancia con nuestros resultados, tanto Helge et al. (22) como Geerling et al. (18), quienes reprodujeron el estudio de Wade et al. (43), no observaron una relación entre RER durante el ejercicio y la adiposidad corporal en varones desentrenados, saludables.
En nuestro estudio y en los estudios anteriores (1, 42) existe una relación consistente entre las concentraciones de FFA circulantes y RER (Tablas 2 y 3). Havel et al. (20) demostraron que la mayoría de la energía que se aporta al músculo esquelético en reposo proviene de los FFAs plasmáticos. Además, el recambio o turnover de FFA plasmáticos en reposo está principalmente controlado por la disponibilidad de sustratos (4) y depende mucho de la ingesta dietaria reciente (29). Por lo tanto, la ingesta previa de alimentos puede alterar la utilización de combustibles modificando la disponibilidad de los sustratos (7). Esta relación entre la ingesta dietaria de grasas en los 3 días previos a la medición de RER fue demostrada en el presente estudio (Tablas 2 y 3) y el estudio de Toubro et al. (42).
Determinantes de la RER en Ayuna Durante el Ejercicio en Estado Estable a Corto Plazo
Otro hallazgo importante de este estudio fue que los determinantes más importantes de RER en ayunas en reposo eran diferentes de los determinantes observados durante el ejercicio. Además, a medida que aumentaba la intensidad del ejercicio, cambiaban as variables utilizadas en los modelos para estimar la RER en las diferentes intensidades de ejercicio, o cambiaba la contribución relativa de las mismas al modelo. En reposo, la composición de fibras musculares fue el determinante más importante de RER en ayuno, pero no contribuyó significativamente con la RER del ejercicio. Durante el ejercicio de baja intensidad (25% Wmax), las concentraciones de sustratos presentes en la sangre fueron los principales determinantes de RER, mientras que durante el ejercicio de intensidad moderada (50% Wmax), las concentraciones de sustratos en el músculo y las actividades enzimáticas se volvieron las variables más importantes para estimar la utilización de sustratos. En los ejercicios de alta intensidad (70% Wmax), la concentración plasmática de lactato fue el determinante mas significativo de RER.
El contenido de glucógeno muscular fue un determinante importante de la utilización de sustratos en la condición de ayuno tanto en reposo como durante el ejercicio. Un aspecto que puede parecer sorprendente es que esta asociación se observó en reposo y durante el ejercicio de baja intensidad (25% Wmax) (Tablas 3 y 4). Romijn et al. (37) observaron que, al 25% del VO2max y en una condición de ayuno en los sujetos entrenados, la mayoría de la energía ara aportada por los FFA y glucosa plasmáticos, con pequeño o ningún aporte del glucógeno muscular. Sin embargo, Friedlander y colaboradores (15, 16) observaron que los CHO explicaban >50% del gasto energético total durante el ejercicio a 45% VO2max en varones y mujeres desentrenados que fueron evaluados 2-3 h después de ingerir una comida estandarizada (448 kcal) que contenía 80g de CHO. Por consiguiente parecería que, independientemente de la intensidad del ejercicio, la oxidación de sustratos puede ser regulada por la disponibilidad de los mismos. De hecho, estudios anteriores (5, 44) han demostrado que la utilización de glucógeno muscular está determinada, en parte, por el contenido de glucógeno muscular al comienzo del ejercicio.
El volumen de entrenamiento también fue un determinante importante de RER, tanto en reposo como durante el ejercicio (Tablas 3-6). Esto coincide con numerosos estudios (3, 8, 28) que han demostrado mayores tasas de oxidación de grasas (y menores tasas de oxidación de CHO) en sujetos entrenados vs sujetos no entrenados, incluso durante ejercicios realizados a la misma intensidad relativa.
Como mencionamos previamente, las concentraciones de sustratos transportados por la sangre particularmente, las concentraciones séricas de FFA, se comportaron como determinantes importantes de RER durante el ejercicio de baja intensidad (25% Wmax). Sin embargo, a medida que la intensidad del ejercicio se incrementaba a 50% Wmax, el peso relativo de la concentración sérica de FFA disminuyó y el contenido de triacilglicéridos en el músculo se volvió un determinante más importante de RER (Tablas 4 y 5). Estos resultados son consistentes con los resultados de Romijn et al. (37) quiénes observaron que la lipólisis periférica (y probablemente la oxidación) era mayor durante el ejercicio de baja intensidad (25% del VO2max) y disminuía progresivamente a medida que la intensidad del ejercicio aumentaba. Mas aún, los autores demostraron que el contenido intramuscular de triacilglicéridos, medido indirectamente con RER y marcadores isotópicos estables, no contribuyo significativamente con la producción de energía a 25% VO2max (10% del total de grasas oxidadas), mientras que a 65% de VO2max, el contenido intramuscular de triacilglicéridos explicó el 50% de la oxidación total de grasas.
No sólo fueron las concentraciones intramusculares de sustrato los determinantes importantes de RER durante el ejercicio, si no que también las actividades enzimáticas musculares contribuyeron significativamente con el modelo de estimación de RER, sobre todo durante el ejercicio de intensidad moderada (50% Wmax). La actividad de CS presentó una correlación inversa con RER al 50% Wmax (r = 0,30, P = 0,026, Tabla 2) y contribuyó significativamente con los modelos que mejor estimaban la RER a 25 y 50% de Wmax (Tablas 4 y 5). Ni Zurlo et al. (47) ni Helge et al. (22) observaron esta asociación, muy probablemente porque sus sujetos no estaban entrenados y tenían menores actividades de CS con un intervalo de valores más pequeño que el de nuestros sujetos. Por otra parte, Zurlo et al. demostraron una relación inversa entre la 24hRER y la actividad de β-HAD (r = -0,75, P = 0,002, Ref. 38) algo que no coincide con nuestros datos (r = 0,008, P = 0,952). Esta diferencia en los resultados no puede ser explicada por las diferencias en el intervalo de valores, pero puede estar relacionada con diferencias metodológicas o con posibles factores de confusión que se mencionan en la Tabla 7.
Nosotros también demostramos una relación positiva entre RER y HK/CS y PFK/CS durante el ejercicio de moderada intensidad (Tabla 2). Estas proporciones nos dan una indicación del flujo glucolítico, que podría haber atenuado la oxidación de ácidos grasos (39), posiblemente controlando la tasa de transporte de ácidos grasos hacia el interior de las mitocondrias (38).
Durante el ejercicio de alta intensidad (70% Wmax), la concentración plasmática de lactato fue el determinante más importante de RER en la ecuación de regresión (Tabla 6). Durante el ejercicio de esta intensidad, se incrementó la oxidación de CHO y utilización de glucógeno muscular y los CHO aportaron más de dos-tercios de la energía necesaria para el ejercicio (para una revisión más detallada, ver la Referencia nº 9). Esto produce tasas aceleradas de producción de lactato en el plasma lo que puede limitar la lipólisis (27). Es más, durante el ejercicio de alta intensidad, la aparición de FFA en el plasma puede disminuir también por la incorporación de FFA al tejido adiposo debido al menor flujo de sangre (23).
RER en Reposo como Determinante de la RER durante el Ejercicio
En este estudio nosotros también observamos que la RER en reposo, independientemente de todos los otros factores, era un buen estimador de RER durante los ejercicios de baja y moderada intensidad [r = 0,60 y r = 0,44 a 25 y 50% Wmax (P <0,0001), respectivamente]. Por consiguiente, a medida que la intensidad del ejercicio aumenta desde el reposo hacia los ejercicios de intensidad creciente, la RER del ejercicio a corto plazo de individuos con valores bajos de RER en reposo aumentará, pero permanecerá relativamente baja en comparación con la de aquellos individuos con valores elevados de RER en reposo que aumentará aún más. Cuando la RER en reposo fue incluida en la ecuación de regresión para predecir la RER en ejercicios de baja y moderada intensidad, aumentó el ajuste de los modelos (Tablas 4 y 5). Esto tiene implicaciones importantes para las investigaciones sobre el metabolismo del ejercicio porque primero, aporta una herramienta útil para estimar fácilmente la utilización relativa de sustratos en atletas, sin requerir que los atletas sean estudiados durante el ejercicio. Segundo, en estudios en donde la utilización de sustratos durante el ejercicio está bajo investigación, puede ser importante considerar la RER en ayuno inicial individual, porque puede impactar en la interpretación de estos estudios.
Diferencias entre Sexos
En el presente estudio, no se observó ninguna diferencia en RER en ayuno entre los varones y mujeres ni en reposo ni durante el ejercicio (Figura 1). Cuando el género se introdujo en el modelo como una variable para predecir RER en reposo y durante el ejercicio, no contribuyó significativamente con el modelo ni alteró su interpretación. Estos resultados son contrarios a los resultados de Horton et al. (25), Friedlander y colegas (14, 16) y Tarnopolsky et al. (41), quienes observaron que las mujeres evaluadas en la fase folicular media del ciclo menstrual presentaron una tendencia a oxidar más grasas durante el ejercicio a la misma intensidad relativa que los hombres. Aunque no se observaron diferencias significativas en el entrenamiento medio por día entre los varones y mujeres en el estudio presente (13,5 vs. 14,6 METS/día, respectivamente), se observó una diferencia significativa en su ingesta dietaria. La ingesta de energía y el % de ingesta dietaria de grasas fueron significativamente más altas y el % de ingesta de CHO fue significativamente menor, en los varones que en las mujeres (12,642 ± 3,247 vs. 8,377 ± 2,654 kJ, 32,6 ± 6,5 vs. 27,0 ± 9,0% de grasas, y 45,6 ± 6,9 vs. 53,4 ± 9,2% CHO, para varones y mujeres, respectivamente, P<0,01). En contraste, en los estudios de Tarnopolsky et al. (41) y Horton et al. (25), la ingesta dietaria de los sujetos se controló durante 2-3 días antes de los tests. Aunque esta diferencia en la ingesta de CHO no produjo diferencias en el contenido de glucógeno muscular de los varones y mujeres (427 ± 126 vs. 436 ±116 mmol/kg peso seco, respectivamente), podría haber enmascarado cualquier diferencia potencial entre los sexos en la utilización de sustratos. De hecho, las mujeres tenían una proporción significativamente mayor de fibras musculares de tipo I (lentas-oxidativas) que los varones (58,4 ± 10,0 vs. 49,6 ± 12,7%, respectivamente, P=0,03), tal como observamos previamente (34). Sin embargo, estos datos deben ser interpretados con cautela debido al número pequeño de mujeres que participaron en este estudio.
Además, en las mujeres no se controló la fase del ciclo menstrual, lo que puede haber influido en la interpretación de las mediciones de RER. Sin embargo, los resultados de las investigaciones existentes, en los cuales se analizó si las fluctuaciones cíclicas de las hormonas sexuales femeninas afectaban la utilización de sustratos durante el ejercicio, no arrojan conclusiones claras (11, 19, 29). Mas aún, Kanaley et al. (29) observaron que la amenorrea no influyó en la utilización de sustratos durante el ejercicio prolongado, en comparación con atletas eumenorreicas evaluadas en la fase folicular temprana y tardía, y en la fase lútea media de su ciclo menstrual.
En el presente estudio, nosotros encontramos una gran variabilidad en la utilización de sustratos en un grupo de ciclistas entrenados que tenían capacidad de rendimiento por encima del promedio. Sin embargo, no se sabe si esta variabilidad en RER, medida durante el ejercicio en estado estable a corto plazo, persistirá por ejemplo durante ejercicios más prolongados, en condiciones de buena alimentación y de disponibilidad baja de glucógeno o cuando se ingieren CHO durante el ejercicio.
Los determinantes más importantes de esta variabilidad y/o su contribución relativa a la utilización de sustratos fueron diferentes en reposo y durante los ejercicios de intensidades diferentes. Sin embargo, en este estudio el entrenamiento, la ingesta dietaria y el consecuente contenido de glucógeno muscular y sustratos circulantes, estimaron consistentemente la RER en reposo y durante el ejercicio. Estos resultados sugieren que, manipulando el entrenamiento y/o la ingesta dietaria, es posible modificar la capacidad de un atleta de alterar la utilización de sustrato durante el ejercicio en estado estable. Además, la RER en reposo se comportó como un determinante independiente significativo de RER a intensidades bajas y moderadas. Este hallazgo puede tener implicaciones importantes para las investigaciones sobre metabolismo, tanto como una herramienta para estimar la utilización relativa de sustratos como para interpretar los resultados de estudios que analicen la utilización relativa de sustratos en respuesta a los diferentes estímulos; sin embargo, es necesario realizar investigaciones adicionales.
Agradecimientos
Agradecemos a todos los sujetos que tan gustosamente participaron en este estudio. Agradecemos a Judy Belonje por su ayuda técnica especializada.
Este estudio fue subsidiado por la Fundación Nacional de Investigaciones de Sudáfrica, el Consejo de Investigaciones Médicas de Sudáfrica, el subsidio Nellie Atkinson y Harry Crossley Staff Research de la Universidad de Ciudad del Cabo y por Alimentos Bromor. Un subsidio adicional fue otorgado por el Programa de Tecnología y Recursos Humanos para la Industria.
Referencias
1. Astrup A, Buemann B, Christensen NJ, Madsen J, Gluud C, Bennett P, and Svenstrup B (1992). The contribution of body composition, substrates and hormones to the variability in energy expenditure and substrate utilization in premenopausal women. J Clin Endocrinol Metab 74: 279286,
2. Bass A, Brdiczka D, Eyer P, Hofer S, and Pette C (1969). Metabolic differentiation of distinct muscle types at the level of enzymatic organisation. Eur J Biochem 10: 198206
3. Bergman BC and Brooks GA (1999). Respiratory gas-exchange ratios during graded exercise in fed and fasted trained and untrained men. J Appl Physiol 86: 479487
4. Bonadonna RC, Groop LC, Zych K, Shank M, and De-Fronzo RA (1990). Dose-dependent effect of insulin on plasma free fatty acid turnover and oxidation in humans. Am J Physiol Endocrinol Metab 259: E736E750
5. Bosch AN, Dennis SC, and Noakes TD (1993). Influence of carbohydrate loading on fuel substrate turnover and oxidation during prolonged exercise. J Appl Physiol 74: 19211927
6. Brooks GA and Mercier J (1994). Balance of carbohydrate and lipid utilization during exercise: the crossover concept. J Appl Physiol 76: 22532261
7. Coggan AR and Mendenhall LA (1992). Effect of diet on substrate metabolism during exercise. In: Perspectives in Exercise Science and Sports Medicine. Energy Metabolism in Exercise and Sport. edited by DR Lamb and CV Gisolfi. Dubuque, IA: Brown and Benchmark, vol. 5, p. 435464
8. Coggan AR, Raguso CA, Gastaldelli A, Sidossis LS, and Yeckel CW (2000). Fat metabolism during high-intensity exercise in endurance-trained and untrained men. Metabolism 49: 122128
9. Coyle EF (1995). Substrate utilization during exercise in active people. Am J Clin Nutr 61: 968S979S
10. Crabtree B and Newsholme EA (1972). The activities of lipases and carnitine palmitoyl transferase in muscles from vertebrates and invertebrates. Biochem J 130: 697705
11. Dombovy ML, Bonekat HW, Williams TJ, and Staats BA (1987). Exercise performance and ventilatory response in the menstrual cycle. Med Sci Sports Exerc 19: 111117
12. Dubowitz V (1985). Muscle Biopsy: A Practical Approach. Eastbourne, England: Bailliere Tindall
13. Durnin JVGA and Womersley J (1974). Body fat assessed from total body density and its estimation from skinfold thickness measurement on 481 men and women aged 1672 years. Br J Nutr 32: 7797
14. Friedlander AL, Casazza GA, Horning MA, Buddinger TF, and Brooks GA (1998). Effects of exercise intensity and training on lipid metabolism in young women. Am J Physiol Endocrinol Metab 275: E853E863
15. Friedlander AL, Casazza GA, Horning MA, Huie MJ, and Brooks GA (1997). Training-induced alterations of glucose flux in men. J Appl Physiol 82: 13601369
16. Friedlander AL, Casazza GA, Horning MA, Huie MJ, Piacentini MF, Trimmer JK, and Brooks GA (1998). Training-inducedalterations of carbohydrate metabolism in women: women respond differently from men. J Appl Physiol 85: 11751186
17. Galbo H, Holst JJ, and Christensen NJ (1979). The effect of different diets and of insulin on the hormonal response to prolonged exercise. Acta Physiol Scand 107: 1932
18. Geerling BJ, Alles MS, Murgatroyd PR, Goldberg GR, Harding M, and Prentice AM (1994). Fatness in relation to substrate oxidation during exercise. Int J Obes 18: 453459
19. Hackney AC, McCracken-Compton MA, and Ainsworth B (1994). Substrate responses to submaximal exercise in the midfollicular and midluteal phases of the menstrual cycle. Int J Sport Nutr 4: 299308
20. Havel RJ, Naimark A, and Borchgrevink CF (1963). Turnover rate and oxidation of free fatty acids of blood plasma in man during exercise: studies during continuous infusion of palmitate-1-C-14. J Clin Invest 42: 10541063
21. Hawley JA and Noakes TD (1992). Peak power output predicts maximal oxygen uptake and performance time in trained cyclists. Eur J Appl Physiol Occup Physiol 65: 798
22. Helge JW, Fraser AM, Kriketos AD, Jenkins AB, Calvert GD, Ayre KJ, and Storlien LH (1999). Interrelationships between muscle fibre type, substrate oxidation and body fat. Int J Obes: 986991
23. Hodgetts V, Coppack SW, Frayn KN, and Hockaday TD (1991). Factors controlling fat mobilization from human subcutaneous adipose tissue during exercise. J Appl Physiol 71: 445451
24. Horton TJ, Pagliassotti MJ, Hobbs K, and Hill JO (1998). Fuel metabolism in men and women during and after long-duration exercise. J Appl Physiol 85: 18231832
25. Hultman, E (1995). Fuel selection, muscle fibre. Proc Nutr Soc 54: 107121
26. Issekutz B Jr, Shaw WAS, and Issekutz TB (1975). Effect of lactate on FFA and glycerol turnover in resting and exercising dogs. J Appl Physiol 39: 349353
27. Jeukendrup AE, Mensink M, Saris WH, and Wagenmakers AJ (1997). Exogenous glucose oxidation during exercise in endurancetrained and untrained subjects. J Appl Physiol 82: 835840
28. Kanaley JA, Boileau RA, Bahr JA, Misner JE, and Nelson RA (1992). Substrate oxidation and GH responses to exercise are independent of menstrual phase and status. Med Sci Sports Exerc 24: 873880
29. Kiens B and Richter EA (1996). Types of carbohydrate in an ordinary diet affect insulin action and muscle substrates in humans. Am J Clin Nutr 63: 4753
30. Klitgaard H, Bergman O, Betta R, Salviati G, Schiaffino S, Clausen T, and Saltin B. (1990). Co-existence of myosin heavy chain 1 and 2a isoforms in human skeletal muscle fibres with endurance training. Pflu¨ gers Arch 416: 470472
31. Ling K, Marcus F, and Lardy H (1965). Purification and some properties of rabbit skeletal muscle phosphofructokinase. J Biol Chem 240: 18931899
32. McNeill G, Bruce AC, Ralph A, and James WPT (1988). Interindividual differences in fasting nutrient oxidation and the influence of diet composition. Int J Obes 12: 455463
33. Nygaard E. (1988). Skeletal muscle fibre characteristics in young wome. Acta Physiol Scand 112: 299304
34. Passonneau JV and Lauderdale VR (1999). A comparison of three methods of glycogen measurement in tissues. Anal Biochem 60:405412
35. Rauch HGL, Hawley JA, Woodey M, Noakes TD, and Dennis SC (1999). Effects of ingesting a sports bar versus glucose polymer on substrate utilization and ultra-endurance performance. Int J Sports Med 20: 252257
36. Romijn JA, Coyle EF, Sidossis LS, Gastaldelli A, Horowitz JF, Endert E, and Wolfe RR (1993). Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am J Physiol Endocrinol Metab 265: E380E391
37. Sidossis LS, Stuart CA, Shulman GI, Lopaschuk GD, and Wolfe RR (1996). Glucose plus insulin regulate fat oxidation by controlling the rate of fatty acid entry into the mitochondria. J Clin Invest 98: 22442250
38. Sidossis LS and Wolfe RR (1996). Glucose and insulin-induced inhibition of fatty acid oxidation: the glucose-fatty acid cycle reversed. Am J Physiol Endocrinol Metab 270: E733E738
39. Srere PA. (1969). Citrate synthase.. Methods Enzymol 13: 311
40. Tarnopolsky LJ, MacDougall JD, Atkinson SA, Tarnopolsky MA, and Sutton JR (1990). Gender differences in substrate for endurance exercise. J Appl Physiol 68: 302308
41. Toubro S, Sorensen TIA, Hindsberger C, Christensen NJ, and Astrup A (1998). Twenty-four-hour respiratory quotient: the role of diet and familial resemblance. J Clin Endocrinol Metab 83: 27582764
42. Wade AJ, Marbut MM, and Round JM (1990). Muscle fibre type and aetiology of obesity. Lancet 335: 805808
43. Weltan SM, Bosch AN, Dennis SC, and Noakes TD (1998). Influence of muscle glycogen content on metabolic regulation. Am J Physiol Endocrinol Metab 274: E72E82
44. Weyer C, Snitker S, Rising R, Bogardus C, and Ravussin E (1999). Determinants of energy expenditure and fuel utilization in man: effects of body composition, age, sex, ethnicity and glucose tolerance in 916 subjects. Int J Obes 23: 715722
45. Zurlo F, Nemeth PM, Choksi RM, Sesodia S, and Ravussin E (1994). Whole-body energy metabolism and skeletal muscle biochemical characteristics. Metabolism 43: 481486
46. Zurlo F, Lillioja S, Esposito-Del Puente A, Nyomba BL, Raz I, Saad MF, Swinburn BA, Knowler WC, Bogardus C, and Ravussin E (1990). Low ratio of fat to carbohydrate oxidation as predictor of weight gain: study of 24-h RQ. Am J Physiol Endocrinol Metab 259: E650E657
Cita Original
Goedecke, Julia H., Alan St Clair Gibson, Liesl Grobler, Malcolm Collins, Timothy D. Noakes, and Estelle V. Lambert. Determinants of the variability in respiratory exchange ratio at rest and during exercise in trained athletes. Am J Physiol Endocrinol Metab 279: E1325-E1334, 2000.
Cita en PubliCE
Timothy D Noakes, Julia H Goedecke, Liesl Grobler, Malcolm Collins, Estelle V Lambert Alan St Clair Gibson (2000). Determinantes de la Variabilidad de la Tasa de Intercambio Respiratorio en Reposo y Durante el Ejercicio en Atletas Entrenados. .https://g-se.com/determinantes-de-la-variabilidad-de-la-tasa-de-intercambio-respiratorio-en-reposo-y-durante-el-ejercicio-en-atletas-entrenados-1356-sa-L57cfb2720279c