Demandas Metabólicas del Entrenamiento Intervalado Aeróbico de Alta Intensidad en Ciclistas de Competición
John A Hawley, David T Martin, Nigel K Stepto, Keiran E Fallon0 Department of Physiology and Applied Nutrition, Australian Institute of Sport, Belconnen 2616, Australia.
Resumen
Objetivo: Investigar las demandas metabólicas de una sola sesión de entrenamiento intervalado aeróbico de alta intensidad en ciclistas de competición altamente entrenados en resistencia. Métodos: Siete ciclistas [consumo de oxígeno máximo (VO2max) 5,14 ± 0,23 L.min-1, Media±DS] realizaron 8 repeticiones de trabajo de 5 min a 86 ± 2% de VO2max con 60 s de recuperación. Inmediatamente antes y después de la sesión de entrenamiento se realizaron biopsias del músculo vasto lateral (vastus lateralis), y se obtuvieron muestras del intercambio de gases pulmonares y de sangre venosa en intervalos regulares a lo largo del ejercicio. Resultados: La concentración de glucógeno muscular disminuyó de 501 ± 91 a 243 ±51 mmol·kg-1 de peso seco (P <0,01). Durante todo el ejercicio se mantuvieron elevadas tasas de oxidación de carbohidratos totales (340 µmol·kg-1 min-1), mientras que la oxidación de grasas aumentó de 16 ± 8 durante la primera repetición de trabajo a 25 ± 13 µmol·kg-1·min-1 durante la séptima repetición (P <0,05). La concentración de lactato sanguíneo se mantuvo entre 5 y 6 mM a lo largo del ejercicio, mientras que la concentración de lactato muscular aumentó de 6 ± 1 en reposo a 32 ± 12 µmol·kg-1 d.m. inmediatamente después de la sesión de entrenamiento (P <0,01). Aunque el pH del músculo disminuyó de 7,09 ± 0,06 en reposo a 7,01 ± 0,03 al final de la sesión (P <0,01), el pH de la sangre fue similar después de las primera y séptima repetición de trabajo (7,34). La saturación arterial de oxígeno (% SPO2) disminuyó a 95,6 ± 1% durante la primera repetición y permaneció en 94% a lo largo del ejercicio. Los intervalos de 60-s de descanso fueron adecuados para restaurar la % SPO2 a 97%. Conclusión: Los ciclistas altamente entrenados pueden mantener elevadas producciones de potencia aeróbica en estado estable que estén asociadas con altas tasas de glucógenolisis y gasto de energía total similares a las experimentadas durante una carrera de competición de 60 min.
Palabras Clave: ácido-base, carbohidratos, glucólisis, lactato
INTRODUCCION
Las características fisiológicas y de rendimiento de los ciclistas de competición han sido ampliamente documentadas (para mas información ver las referencias 3,11). Sin embargo, la literatura científica sobre los efectos puntuales de procedimientos de entrenamiento específicos sobre el rendimiento de individuos previamente entrenados es escasa: los científicos deportivos han encontrado dificultades para persuadir a atletas de élite para que realicen experimentos con sus regimenes de entrenamiento (11). Una forma de entrenamiento que ha sido utilizada por atletas de resistencia durante varias décadas y que es muy reconocida por los científicos del deporte, es el entrenamiento intervalado. Tal entrenamiento normalmente se inicia después de que un atleta ha completado un gran volumen de trabajo de resistencia y generalmente ha consistido en realizar varias repeticiones de ejercicio aeróbico sostenido de 3-15 min de duración, alternadas con intervalos de descanso más cortos (60-90 s) de actividad a un ritmo más lento. El objetivo de tal entrenamiento es exponer los sistemas fisiológicos de potencia al ejercicio sostenido a una intensidad (o esfuerzo) qué corresponda al mayor “ritmo en estado estable” actual del atleta (11), también llamado "umbral del lactato" individual (17). Los beneficios atribuidos al entrenamiento intervalado aeróbico incluyen una mejor cinética del lactato, estímulo de patrones neurológicos específicos de reclutamiento de fibras musculares necesarias para el ritmo de una carrera, aumento en la resistencia a la fatiga y mejor rendimiento atlético (11, 18, 32, 33).
Aunque el entrenamiento intervalado ha sido un elemento básico en el acondicionamiento físico desde el comienzo del siglo, existen pocos estudios sobre los efectos del entrenamiento intervalado sobre el rendimiento de atletas competitivos altamente entrenados (1, 11, 18, 32, 33, 34). De hecho, es posible argumentar que, en la actualidad, los científicos deportivos están haciendo recomendaciones sobre el papel de diferentes regímenes de entrenamiento en la preparación de atletas, sin el conocimiento preciso de las demandas metabólicas asociadas con tales intervenciones. Debido a la escasez de bibliografía en esta área, recientemente abordamos una serie de investigaciones en el laboratorio utilizando ciclistas altamente entrenados para estudiarar la evolución de los cambios en el rendimiento deportivo en respuesta a los diferentes programas de entrenamiento intervalado de alta intensidad (11, 18, 32) y analizar las adaptaciones metabólicas (33) y musculares (34) crónicas que ocurren en respuesta a dicho entrenamiento. En el presente estudio, nosotros ampliamos esta información determinando las demandas metabólicas de una sola sesión de entrenamiento intervalado aeróbico de alta intensidad en ciclistas competitivos.
MÉTODOS
Sujetos
Siete ciclistas altamente entrenados (Tabla 1) que pedaleaban 368 ±141 km-sem-1 (Media±DS) en Canberra, a una altitud de 588 m, y que no habían realizado ningún entrenamiento intervalado de alta-intensidad en las 4 semanas previas a la investigación, fueron reclutados para participar en este estudio. Todos los sujetos habían participado en estudios previos y estaban familiarizados con los procedimientos de evaluación y fueron completamente informados de los riesgos potenciales asociados al estudio antes de proporcionar su consentimiento informado por escrito. Este estudio fue aprobado por el Comité de Ética de Investigación Humana de la Universidad de RMIT y el Comité de Ética del Instituto Australiano del Deporte. Dados los objetivos de esta investigación no se utilizó ningún grupo comparativo o “control”: estudios piloto revelaron que los sujetos desentrenados eran incapaces de pedalear más de 90s en la misma producción de potencia absoluta sostenida por nuestros ciclistas altamente entrenados durante 40 min y que, a la misma intensidad de ejercicio relativa, ellos no podrían alcanzar las condiciones de estado estable necesarias para obtener mediciones válidas de oxidación de sustratos (28). Puntualmente, nosotros hemos informado previamente los efectos crónicos de entrenamiento intervalado de alta intensidad (6 sesiones) sobre los cambios en la oxidación de sustratos (33).
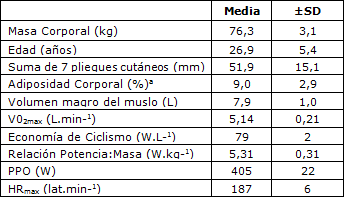
Tabla 1. Características de los Sujetos. V02max= Consumo de oxígeno máximo; HRmax = Frecuencia cardíaca máxima; PPO= producción de Potencia Máxima Sostenida. a Estimada a partir de las ecuaciones de Withers et al. (35). Todos los valores se expresan en forma de Media ± DS. n = 7.
Evaluación Preliminar
En su primera visita al laboratorio, los sujetos fueron pesados y se les realizó la medición de los parámetros antropométricos para la suma de siete pliegues cutáneos (bíceps, tríceps, subescapular, abdominal, supra-espinal, medio-muslo y gastrocnemio medio) y el volumen magro del muslo (16). Luego, después de una entrada en calor en un ritmo autoseleccionado (30 min) los participantes realizaron un test de ciclismo máximo, incremental hasta el agotamiento en una bicicleta ergométrica con freno electrónico (Lode, Gronigen, Países Bajos). El protocolo de prueba se describió previamente con detalle (12). Para evaluar la exactitud y confiabilidad de la bicicleta ergométrica se utilizó un equipo de calibración de primeros principios. La producción de potencia esperada (W) se situó dentro de ± 2% de la potencia real de 200 a 800 W cuando la frecuencia del pedal estaba entre 90 y 140 rev·min-1. A lo largo de la prueba máxima y del tratamiento experimental que se describe a continuación, los sujetos inspiraron el aire a través de una válvula de Hans Rudolph bidireccional acoplada a un sistema de análisis de gases automatizado con una bolsa de Douglas fabricada a pedido (Instituto Australiano del Deporte, ACT, Australia) que incorporó analizadores de O2 y CO2 (Ametek N-22 electrochemical O2 sensor, model S3A, and Ametek P-61B infrared CO2 sensor, Applied Electrochemistry, Ametek Instruments, Pittsburgh, PA) y dos gasómetros de Tissot (Warren E. Collins Inc., Braintree, MA) conectados a una computadora personal IBM por medio de un Codificador Óptico Rotatorio (RS 341-597, Berna, Suiza) que calculó la tasa de O2 consumo (VO2), producción de CO2 (VCO2), el ventilación por minuto (VESTPD) y la tasa de intercambio respiratorio (RER) cada 30s a partir de ecuaciones convencionales. Antes de realizar el test de intensidad máxima y todos las pruebas experimentales, se calibraron los analizadores con gases α con volumen conocido de O2 y CO2. Antes y después de realizar el estudio se utilizó un calibrador automático de alta-capacidad para calorimetría indirecta de circuito abierto para verificar simultáneamente los analizadores de gases, dispositivos de volumen y software del sistema realizado a pedido (9). Este dispositivo puede calibrar volúmenes de ventilación altos (100 L.min-1) medidos cuando los atletas altamente entrenados realizan trabajo durante períodos sostenidos de ejercicios de intensidad elevada (80-90% del consumo de oxígeno máximo (VO2max).
El VO2max se definió como el mayor consumo de O2 que un sujeto alcanzó durante dos períodos de muestreo consecutivos de 30s. La producción de potencia máxima sostenida (PPO) se calculó a partir de la última tasa de trabajo completada, más la fracción de tiempo gastado en la tasa de trabajo final no completada multiplicada por 25 W (12). Los resultados del test máximo se utilizaron para determinar la producción de potencia que correspondía al 85% del VO2max de cada sujeto que sería utilizada en la sesión de entrenamiento intervalado aeróbico de alta intensidad que se describe posteriormente.
Entrenamiento y Control de la Dieta
Veinticuatro horas antes de la prueba experimental se controló el entrenamiento y el estado nutricional de los sujetos para estandarizar las reservas de glucógeno en el músculo y en el hígado. El día previo a la prueba, todos los sujetos arribaron al laboratorio entre las 07:00 y 08:00 h y pedalearon durante 60-min a 50% de PPO (60% VO2max). Luego se les proporcionó una dieta estándar de 50 kcal·kg-1 de masa corporal (BM), compuesta de 63% de carbohidratos (8 g·kg-1 BM), 20% grasa y 17% proteínas, para ser consumida a lo largo de las próximas 24 h. Durante este tiempo, los sujetos se abstuvieron de realizar cualquier actividad de entrenamiento adicional.
Prueba Experimental
El día de la prueba, los sujetos arribaron al laboratorio entre las 07:00 y 08:00 h después de un ayuno de toda la noche. Fueron pesados y luego descansaron sentados en silencio durante 10-15 min. Seis de éstos sujetos de élite expresaron su conformidad de sufrir un máximo de dos biopsias musculares. De acuerdo con esto, nosotros decidimos muestrear el músculo inmediatamente antes y después del ejercicio para evaluar las demandas metabólicas de la sesión de entrenamiento intervalado completa (que se describe posteriormente), en lugar de la respuesta a una sola de trabajo. Después de colocar la anestesia local a la piel, tejido hipodérmico y fascia del vasto lateral, se realizó una biopsia en reposo aplicando succión con una aguja de Bergstrom de 6 mm. Se extrajeron aproximadamente 150 mg de músculo que rápidamente (18 ± 6 s) fueron congelados en nitrógeno líquido. Las muestras fueron almacenadas a -80°C hasta el análisis posterior. Inmediatamente después de esta biopsia, se midió la temperatura del músculo mediante un termómetro aguja (YSI 525, Yellow Springs Instruments, Yellow Springs, OH) que fue insertado a una profundidad de 4 centímetros a través de la misma incisión (7). En este momento, otro sitio en la misma pierna (5 centímetros de distancia) fue preparado para la segunda biopsia que se realizaría inmediatamente después del ejercicio. Luego se insertó una cánula de Teflón en la vena antecubital del antebrazo para efectuar un muestreo rápido continuo de sangre a través de un robinete estéril. La cánula era regularmente rellenada con una solución salina estéril al 0,9% para mantener la vena abierta.
A continuación los ciclistas subieron a la bicicleta ergométrica y comenzaron una entrada en calor estandarizada que consistió en 2 min a 25%, 50%, y 60% de PPO. Después de un descanso de 2-min durante el cual los sujetos permanecieron en la bicicleta ergométrica, comenzaron la sesión de entrenamiento intervalado que consistió en 8 repeticiones de trabajo de 5 min al 82,5% de PPO (86 ± 2% de VO2max: 334 ± 18 W) con 60 s de recuperación activa (una relación trabajo: descanso de 5:1). Durante la recuperación de 60s entre las repeticiones de trabajo, los sujetos pedaleaban con una carga de trabajo de <100 W. Nosotros informamos previamente que los ciclistas altamente entrenados que realizan seis sesiones de este protocolo de entrenamiento intervalado aumentan significativamente su rendimiento en prueba contrarreloj de 40 km de ciclismo (11, 18, 32, 33). Las condiciones del laboratorio se mantuvieron a 22°C y 46% humedad relativa. Los sujetos fueron refrescados con un ventilador (velocidad del viento de 7 m·s-1) y se les aportó la cantidad de agua que desearan a lo largo del ejercicio.
Durante la primera, tercera, quinta y séptima repetición de trabajo y durante los 60s de recuperación subsiguientes, los sujetos respiraron a través de una boquilla acoplada al sistema analizador de gases automatizado descripto previamente. Además, se recolectaron los datos de intercambio de gases pulmonares de tres sujetos a lo largo de la última serie de trabajo. Las muestras de sangre (10 mL) se tomaron en reposo, inmediatamente después de la entrada en calor y al final de la primera, tercera, cuarta, quinta, séptima y octava repeticiones.
La tasa de oxidación de carbohidratos (CHO) y grasas (g·min-1) en el cuerpo entero se calculó a lo largo de las repeticiones 1, 3, 5, y 7 a partir de VCO2 y VO2 usando los valores de RER del metabolismo no proteico, según las siguientes ecuaciones (24):
Oxidación de CHO = 4,585 VCO2 - 3,226 VO2
Oxidación de grasas = 1,695 VO2 - 1,701 VCO2
Estas ecuaciones se basan en la suposición que VO2 y VCO2 reflejan con precisión el consumo de O2 y producción de CO2 de los tejidos. En los sujetos altamente entrenados como los que participaron en la investigación actual, se ha demostrado previamente que la calorímetría indirecta es un método válido para cuantificar tasas de oxidación de sustratos durante ejercicios extenuantes a 85% del VO2max (28).Las tasas de oxidación de sustratos en g·min-1 fueron transformadas a µmol·kg-1·min-1 para que pudieran ser comparadas con los resultados de estudios anteriores. Las tasas de oxidación de ácidos grasos (FA) (µmol·kg-1·min-1) fueron determinadas transformando la tasa de oxidación de triacilglicéridos expresada en g·min-1, en su equivalente molar asumiendo que el peso molecular medio de los triacilglicéridos humanos es 855,26 g·mol-1 y multiplicándola por la tasa molar de oxidación de triglicérido por tres, porque cada molécula contiene 3 mol de FA. Las tasas de oxidación de CHO (µmol·kg-1·min-1) se determinó convirtiendo la tasa de oxidación de carbohidratos en g·min-1 a su equivalente molar asumiendo que se consumen 6 moles de O2 y se producen 6 moles de CO2 por cada mol oxidado (180 g).
Durante todas las sesiones del ejercicio, la frecuencia cardíaca de los sujetos (HR) fue continuamente monitoreada mediante telemetría (Accurex Plus; Polar Electro OY, Kempele, Finland). La saturación de oxígeno de la hemoglobina (Hb) arterial (% SpO2) se supervisó a lo largo las repeticiones 1, 3, 5 y 7 y la recuperación subsecuente mediante un oxímetro de pulso para la yema de los dedos (modelo U:S 504, Criticare, Waukesha, WI). Previamente se ha demostrado que esta técnica proporciona mediciones válidas de % SpO2 durante el ejercicio de alta intensidad (21). Inmediatamente después de la sesión de entrenamiento, se realizó una segunda biopsia mientras los sujetos permanecían sentados en la bicicleta ergométrica y la muestra fue congelada rápidamente (18 ± 8 s) en nitrógeno líquido. Posteriormente se midió, la temperatura de músculo post-ejercicio tal como se describió previamente.
Análisis
Las muestras de sangre (5 mL) fueron analizadas inmediatamente para determinar la concentración sanguínea de glucosa y lactato, K+, hemoglobina, hematocrito y pH en un analizador de gases sanguíneos ABL serie 700 (Radiometer Medical A/S, Dinamarca). Los cambios en el volumen del plasma fueron estimados mediante las ecuaciones de Dill y Costill (5). Las muestras de músculo congeladas, fueron pesadas, deshidratadas por congelación, pesadas nuevamente y seccionadas libres de sangre y de tejido conjuntivo. Luego estas muestras fueron divididas en dos alícuotas separadas. Una alícuota (1 mg de peso seco [d.m.]) se extrajo en ácido perclórico 3 M (PCA) congelado en nitrógeno líquido, se llevó la temperatura a -8°C durante 20 min y finalmente se colocó a una temperatura de 5°C durante 5 min. Luego en este extracto se realizó el análisis de lactato muscular a través de una técnica enzimática fluorométrica estándar (19). La concentración de glucógeno muscular se evaluó por duplicado en la segunda alícuota según los procedimientos de Passonneau y Lauderdale (23).
El pH del músculo se determinó de la porción de músculo restante congelada (5 mg), usando la técnica modificada de Sahlin (31). La muestra de músculo fue homogeneizada en una solución de homogeneización enfriada con hielo (145 mM KC1, 10 mM NaCl, y 5 mM de iodoacetato de sodio; pH 7) y luego fue analizada usando un sistema de análisis de gases sanguíneos CIBA Corning 278 (CIBA Corning Diagnostic Corps., Medfield, MA).
Análisis Estadísticos
Todos los valores se informan como Media ± SD. El análisis de la concentración muscular de glucógeno y lactato, la temperatura del músculo y pH del músculo antes y después del ejercicio se realizó mediante un test-t de Student de muestras apareadas. Los datos restantes (HR, tasa de oxidación de sustratos, concentración de lactato sanguíneo y concentración de glucosa en la sangre, HCO3 y K+) fueron analizados por medio del ANOVA de mediciones repetidas de una vía. En los casos en que se observara un índice F significativo, los datos se analizaron mediante el test post hoc de Tukey. La significancia estadística se fijó en P <0,05.
RESULTADOS
Intercambio de Ggas Pulmonar, Frecuencia Cardíaca y Tasa de Oxidación de Sustratos
En la Tabla 2 se presentan las mediciones del intercambio de gases pulmonares y las tasas estimadas de oxidación de sustratos del cuerpo entero medidas a lo largo de la primera, tercera, quinta y séptima repeticiones de trabajo de la sesión de entrenamiento intervalado, mientras que en la Figura 1 se presentan las respuestas de VO2 y de HR.
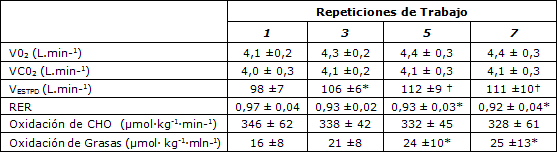
Tabla 2. Mediciones de intercambio de gases pulmonares y tasas de oxidación de sustratos en todo el cuerpo determinadas durante la sesión de entrenamiento intervalado de alta intensidad. VO2=Consumo de oxígeno; VCO2= Producción de dióxido de Carbono; VE,=ventilación; RER=Tasa de intercambio respiratorio; CHO= Carbohidratos. Los valores se presentan en forma de Media ± SD (n = 7). * Presenta diferencias significativas respecto a la repetición 1 (P < 0,05); † Presenta diferencias significativas respecto la repetición 1, (P < 0,001).
A pesar de los valores similares de VO2 y VCO2 a lo largo de la sesión de entrenamiento, la VE aumentó de 98 ± 7 L.min-1 durante la primer repetición de trabajo a 106 ± 6 L.min-1 después de la tercera repetición (P <0,05). En la quinta repetición de trabajo, VE se había incrementado a 112 ± 9 L-min-1 (P <0,001 en comparación con la repetición 1) pero no siguió incrementándose durante la séptima repetición. El tiempo necesario para alcanzar un VO2 en estado estable igual a 4,45 ± 0,25 L-min-1 (promedio de los últimos 3 min de cada repetición de trabajo), fue significativamente mayor durante la primera repetición en comparación con la tercera, quinta y séptima repeticiones de trabajo (185 ± 30 vs 145 ± 35, 130 ± 25 y 130 ± 36 s, respectivamente; P <0,05, Figura 1A). La Figura 1B muestra la respuesta de HR a la sesión de entrenamiento de alta intensidad. La frecuencia cardíaca (HR) a lo largo de la primera repetición de trabajo (156 ± 8 latidos·min-1) fue significativamente más baja que los valores observados en las repeticiones dos a ocho (162 ± 7 latidos·min-1: P<0,05). Como era de esperar, se observó un desplazamiento o drift de la HR gradual hacia arriba, durante el ejercicio a lo largo de la sesión de entrenamiento intervalado. Durante cada recuperación de 1 minuto, la HR disminuyó 35 lat·min-1 (P <0,01). La HR de la recuperación para la repetición 1 fue (125 ± 8 latidos·min-1) y fue significativamente menor que la observada después de todas las otras repeticiones de trabajo (Figura 1B, P<0,05)
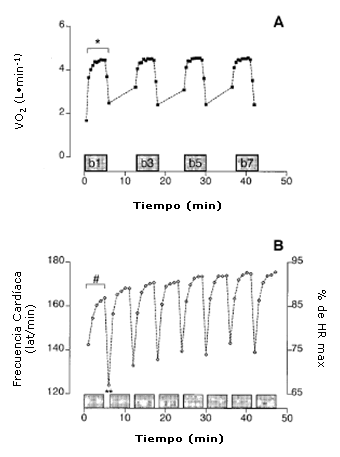
Figura 1. Respuestas del consumo de oxígeno (V02) (A, panel superior) durante la primera, tercera, quinta y séptima repeticiones de trabajo y respuestas de la frecuencia cardíaca (HR) (B, panel inferior) durante cada repetición de trabajo y durante los siguientes 60s de recuperación determinadas durante la sesión de entrenamiento intervalado de alta intensidad. Los valores se expresan en forma de medias(N = 7); * se observaron diferencias significativas respecto a las repeticiones 3, 5, y 7 (P <0,05); * * se observaron diferencias significativas respecto a todas las otras recuperaciones (P <0,05). # Presenta diferencias significativas respecto a todas las otras repeticiones de trabajo (P <0,05).
Los valores de RER disminuyeron progresivamente a lo largo de la sesión de entrenamiento intervalado de 0,97 ± 0,04 durante la primera repetición a 0,92 ± 0,04 en la séptima repetición. En consecuencia, la tasa media de oxidación de CHO disminuyó ligeramente, pero no significativamente, de la primera a la séptima repetición de trabajo (de 346 ± 62 µmol·kg-1·min-1 a 312 ± 58 µmol·kg-1·min-1 mientras que la tasa de oxidación de grasas aumentó (de 16 ± 8 µmol·kg-1·min-1 a 25 ± 13 µmol·kg-1·min-1, P <0,05). Los datos de intercambio de gases pulmonares fueron recolectados de tres sujetos durante la octava repetición de trabajo. Estos datos revelaron que no había ninguna diferencia en VO2, VCO2, VE o RER en comparación con la séptima repetición de trabajo. La tasa de gasto de energía total fue 1252 ± 96 J·kg-1m·min-1.
Parámetros Sanguíneos
La Figura 2 muestra los resultados de las mediciones obtenidas a partir de la sangre venosa, en reposo, inmediatamente después de la entrada en calor y durante la primera, tercera, quinta y séptima repeticiones de trabajo. La concentración de lactato sanguíneo (Panel A) aumentó de 1,0 ± 0,3 mmol-L-1 en reposo a 4,7 ± 1,7 mmol.L-1 (P <0,05) luego de la primera repetición. Después de esto, se mantuvo constante entre 5-6 mmol.L-1 durante las restantes repeticiones del entrenamiento. En contraste, la concentración de HCO3- en la sangre (Panel B) disminuyó de 26,1 ± 2 mmol-L-1 en reposo a 22,2 ±1,1 mmol-L-1 (P <0,05) después de la primera repetición de trabajo. A partir de allí, las concentraciones de HCO3- se mantuvieron en 21 mmol.L-1. El pH sanguíneo (Panel C) en reposo fue de 7,35 ± 0,03 y disminuyó a 7,32 ± 0,02 después de la primera repetición de trabajo (P> 0,05), momento a partir del cual aumentó ligeramente de manera que en la quinta repetición de trabajo se había recuperado hasta valores cercanos a los valores de reposo. La concentración sanguínea de K+ (Panel D) en reposo era 4,0 ± 2 mmol·L-1 y aumentó a 4,8 ± 0,2 mmol·L-1 (P <0,05) al final de la primera repetición de trabajo. La concentración de K+ se incrementó a 5,4 ± 0,4 mmol·L-1 (P <0,05) después de la tercera repetición de trabajo y se mantuvo hasta el fin de la sesión de entrenamiento. La concentración sanguínea de glucosa fue 5,02 ± 0,41 mmol·L-1 en reposo y sólo aumentó a 5,6 ± 0,6 mmol·L-1 después de la séptima repetición de trabajo. Las muestras de sangre tomadas de tres sujetos no revelaron ninguna diferencia en ninguna de las variables de la sangre entre la séptima y octava repetición de trabajo. El volumen plasmático disminuyó 9,8 ± 2,8% con respecto a los valores en reposo inmediatamente después de la entrada en calor y sólo disminuyó un 2% adicional (a 12,1 ± 3,4%) al final del ejercicio.
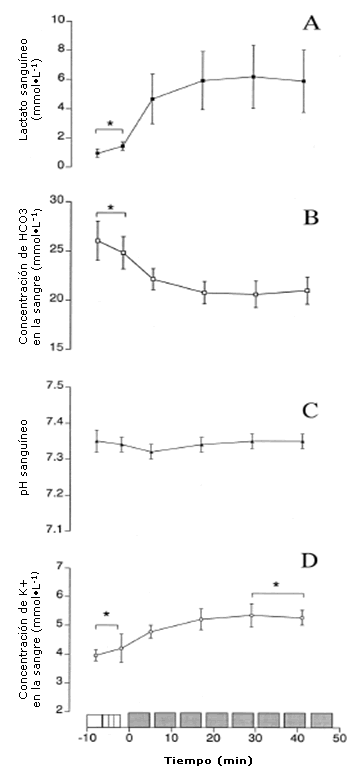
Figura 2. Concentración de lactato en sangre venosa (A), concentración de HCO3 (B), pH sanguíneo (C) y concentración de K+ en la sangre (D) medidos en reposo (·), después de la entrada en calor y durante la primera, tercera, quinta y séptima repeticiones de trabajo de la sesión de entrenamiento intervalado de alta intensidad. Los valores se presentan en forma de Media ± DS (n=7); * se observaron diferencias significativas respecto de la repetición 1 (P <0,05).
Parámetros Musculares
La Figura 3 muestra los datos obtenidos antes y después del ejercicio en las muestras de músculo. La concentración de glucógeno muscular en reposo (Panel A) era 501 ± 91 mmol-kg-1 de masa seca. y disminuyó a 243 ±51 mmol-kg-1 de masa seca después de la sesión de entrenamiento intervalado (P <0,01). La concentración de lactato muscular en reposo (Panel B) y el pH muscular (Panel C) fueron 6,2 ± 0,8 mmol.kg-1 de masa seca y 7,09 ± 0,06, respectivamente. La concentración de lactato muscular aumentó significativamente a 32,7 ± 12,2 mmol.kg-1 de masa seca (P <0,01), mientras que el pH del músculo disminuyó a 7,01±0,03 (P <0,01) inmediatamente luego del final del entrenamiento. Se observó una correlación significativa entre la concentración media de lactato durante el ejercicio y la concentración de lactato post ejercicio (r=0,91, P <0,05).La temperatura muscular (Panel D) en reposo era 33,6 ± 2°C y había aumentado a 36,8 ± 2°C (P <0,001) al final de la sesión de entrenamiento intervalado.
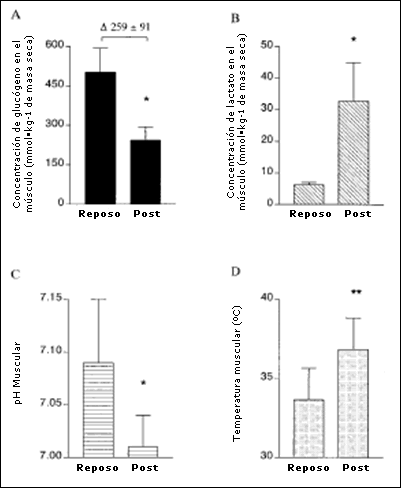
Figura 3. Concentración de glucógeno en el músculo (A), concentración de lactato en el músculo (B), pH del músculo (C) y temperatura del músculo(D), antes y después de la sesión de entrenamiento intervalado de alta intensidad. Los valores se presentan en forma de Media ± DS (N = 6); * significativamente diferente del reposo (P <0,01); * * significativamente diferente del reposo (P <0,001).
% SPO2
La saturación de oxígeno arterial (% SPO2) fue 95,6 ± 1% durante la primer repetición de trabajo y disminuyó a 94 ± 2% (P <0,05) en la tercera, quinta y séptima repeticiones de trabajo. Los intervalos de 60 s de descanso fueron adecuados para restaurar la % SP02 a 97% a lo largo de toda la sesión de entrenamiento intervalado de alta intensidad.
DISCUSIÓN
Las respuestas fisiológicas y metabólicas de individuos altamente entrenados frente a ejercicios de ciclismo de resistencia continuos, prolongados y de moderada intensidad están ampliamente documentadas (3). También hay información sustancial sobre los cambios del músculo de esquelético en la utilización de sustratos después de esprints repetidos máximos de corta duración (<30 s) en carreras de ciclismo y cinta rodante (para la revisión ver la referencia 22). Sin embargo, hay pocos datos que documenten las perturbaciones metabólicas que se producen en respuesta a repeticiones repetidas de ejercicio aeróbico similares en intensidad (es decir, 85% VO2máx) y duración (es decir, 1 h) a las que enfrentan los atletas de competición durante el entrenamiento y las competiciones (33). Aquí, por primera vez, nosotros informamos las demandas metabólicas asociadas con las elevadas tasas de trabajo (340 W) absolutas que los ciclistas de resistencia competitivos pueden sostener durante una sola sesión de entrenamiento intervalado. Nuestras investigaciones previas han determinado los cambios en los modelos de oxidación de sustratos (33), rendimiento (18,32) y en ciertos parámetros musculares (34) que ocurren en respuesta a una intervención crónica (6 sesiones).
El primer hallazgo fue la elevada tasa media de oxidación total de CHO (340 µmol·kg-1·min-1) mantenida a lo largo del ejercicio (Tabla2). Tales tasas de oxidación son similares a las informadas previamente (350 µmol·kg-1·min-1) para los ciclistas altamente entrenados (VO2max 5,2 L·min-1) que pedaleaban a 325 W durante 50 min (10) y probablemente se encuentran cerca del límite superior para las tasas en estado estable de utilización de combustible que es posible mantener durante 1 h. Solo conocemos otras tres investigaciones que informaron tasas en estado estable de utilización de sustratos durante la realización de ejercicio aeróbico de alta intensidad (4, 29, 30). Romijn et al. (30) observaron que durante los últimos 10 min de una serie de trabajo de 30-min a 85% de VO2max, la tasa de oxidación de CHO era 260 µmol·kg-1·min-1. Aunque este valor es aprox 35% menor que la tasa promedio de oxidación de CHO encontrada en el presente estudio, esto probablemente se deba a que los sujetos de nuestro estudio mantuvieron una tasa de trabajo absoluta 25 W más alta, que los sujetos del estudio de Romijn et al. (30). Coyle et al. (4) informaron una tasa media de sólo 170 µmol·kg-1·min-1 para la oxidación total de CHO durante 30 min de ciclismo al 80% del VO2max en un grupo de siete ciclistas de competición (VO2max 4,87 L·min-1) con un "umbral de lactato" alto. Si nosotros suponemos que estos sujetos mantuvieron una tasa de trabajo de 300 W (12), entonces no sólo es una tasa de utilización de sustratos sumamente baja, si no que el valor prevaleciente de RER= 0,85 significa que la contribución de los CHO a la energía fue sólo 52%. En el presente estudio, el valor de RER durante las últimas fases de ejercicio fue 0,92 (74% de energía de CHO) que tiene una gran coincidencia con el valor de 0,91 informado por Romijn et al. (29) para ciclistas igualmente bien entrenados (VO2max 5,04 L-min-1) durante los últimos 10 min de un ejercicio de ciclismo de 30-min al 85% de VO2max. De hecho, en la investigación presente y en el estudio de Romijn et al. (29) no sólo fueron similares las tasas oxidación de CHO y grasas a la misma intensidad relativa de ejercicio, si no que también fueron comparables durante las últimas etapas del ejercicio las tasas absolutas de oxidación de grasas (25 contra 27 µmol·kg-1·min-1) .
Esta ampliamente establecido que la utilización de glucógeno y de glucosa aumentan exponencialmente en función de la intensidad relativa del ejercicio, siendo mayor el aumento en la oxidación de glucógeno que en la oxidación de glucosa en las producciones de potencia más altas (para una revisión ver la referencia 2). En el estudio actual, la cantidad absoluta de glucógeno oxidada durante los 40 min de ejercicio fue 260 mmol.kg-1 de peso seco, lo que equivale a una tasa media de desaparición de glucógeno de 6,5 mmol.kg-1 de peso seco·min-1 en el músculo donde se realizó la biopsia. Como era de esperar, una tasa de utilización de glucógeno así, es algo más alta que la tasa de 4,6 mmol.kg-1 de peso seco.min-1 calculada para los ciclistas entrenados durante una prueba por contrarreloj de 1 hora (13) y considerablemente superior al valor estimado de 4 mmol.kg-1 de peso seco·min-1 calculada de los datos de Coyle et al. (4). En el presente estudio, la utilización de glucógeno explicó el 90% de la oxidación total de CHO. Este porcentaje es ligeramente mayor que la contribución del 85% de la glucógenolisis a la utilización total de CHO estimada por Romijn et al. (29) usando una combinación de marcadores de isótopos estables y calorimetría indirecta y notablemente superior que los 77% informados por Coyle et al. (4) a partir de mediciones directas de desaparición de glucógeno muscular y datos de RER.
Hay varias razones posibles para explicar tal diferencia. La más probable es una sobrestimación de la masa de músculo activo de nuestros sujetos. Las ecuaciones de Katch y Katch (16) asumen que el muslo es un cono truncado que incluye el músculo activo y todos los demás tejidos incluyendo el hueso (fémur), pero excluyendo la grasa subcutánea y la piel. Segundo, nuestras mediciones realizadas en el músculo antes y después del ejercicio representan la desaparición total de glucógeno a lo largo de la sesión de entrenamiento completa, incluyendo la transición desde el reposo e incorporando los 10 min iniciales de ejercicio, momento en el cual, se sabe que la tasa de glucógenolisis es muy el rápida (6). Por otra parte, las estimaciones de utilización de glucógeno muscular efectuadas en el estudio de Romijn et al. (29) fueron realizadas durante las últimas fases de ejercicio, en un momento en el que se esperaría que la contribución del glucógeno muscular sea más baja y la contribución de la oxidación de la glucosa sanguínea sea superior a los requerimientos globales de CHO del ejercicio (2). En el estudio de Coyle et al. (4), tanto la intensidad del ejercicio absoluta como la relativa fueron más bajas que las empleadas en la presente investigación, condiciones que se esperaría que produzcan una mayor utilización de lípidos y una menor tasa de oxidación total de CHO. A pesar de estas diferencias está claro que, incluso en los ciclistas altamente entrenados cuya musculatura se supone que tendrá una mayor capacidad de captación y oxidación de ácidos grasos libres (FFA) plasmáticos y/o triacilglicéridos intramusculares, la selección del sustrato durante el ejercicio de alta intensidad está dominada por la oxidación de CHO (2).
Como observamos previamente, pocos estudios han informado tasas de utilización de sustratos durante los ejercicios aeróbicos de alta intensidad porque se supone que en intensidades por encima del denominado "umbral del lactato" el VCO2 no es completamente una función de la producción del tejido si no que también refleja un agotamiento del pool de bicarbonato, para mantener la homeostasis ácido base. Sin embargo, la calorimetría indirecta puede proporcionar tasas de oxidación de sustratos válidas durante el ejercicio de alta intensidad, en aquellos sujetos entrenados en resistencia que son capaces de mantener elevadas tasas de trabajo absolutas (y relativas) con un equilibrio ácido-base estable, tal como lo refleja la concentración de lactato sanguíneo prevaleciente (28). En el presente estudio, los valores de VCO2 permanecieron estables a lo largo del ejercicio (Tabla 2), mientras que la concentración de lactato en sangre se incrementó a 5 mM después de la primera repetición de trabajo y después de esto se estabilizó durante el resto de la sesión de entrenamiento (Figura 2). Las concentraciones de lactato en sangre son similares al valor en estado estable de 7 mM informados por otros autores para ciclistas de resistencia altamente entrenados durante la realización de ejercicio de alta intensidad (85-88% de VO2max) (4, 30). Las alteraciones en el equilibrio ácido-base fueron más marcadas después de la primera repetición de trabajo (Figura 2). A partir de allí, la acidosis metabólica inicial probablemente fue compensada por un flujo de iones H+ que provocaron una caída inicial en el pH sanguíneo y en la concentración de HCO3- (Figura 2). Una vez que este flujo de iones H+ fue amortiguado por el HCO3-, el pH de la sangre retornó a niveles cercanos a los del reposo durante el resto de las repeticiones del ejercicio, lo que demostró el buen control metabólico en estos ciclistas.
Las mediciones del estado ácido base del músculo obtenidas de las biopsias realizadas inmediatamente al finalizar el ejercicio revelaron una correlación significativa (r 0,91, p <0,05) entre la concentración de lactato en sangre y la concentración prevaleciente de lactato en el músculo, tal como se ha informado previamente (15). Los cambios en la saturación arterial de O2 durante la sesión de entrenamiento de alta intensidad se correspondieron con informes previos, en los cuales se determinó el % SPO2 en los atletas entrenados, capaces mantener tasas metabólicas en estado estable cercanas a 4,5 L-min-1 (27). Típicamente, el %SPO2 disminuye a alrededor de 94% durante ejercicios de alta intensidad de corto plazo (27) y, dependiendo del intervalo de descanso entre las repeticiones de trabajo, se recupera a valores entre 96 y 97%. En el estudio actual, 60s de recuperación activa fueron suficientes para restaurar el %SPO2 a 97% a lo largo de la sesión de ejercicios completa.
Las cinéticas de VO2 en el comienzo de cada repetición de trabajo fueron rápidas (Figura 1A) tomando 3 min para alcanzar el valor alto (4,3-4,4 L.min-1) de consumo de oxígeno en estado estable que nuestros sujetos fueron capaces de mantener durante toda la sesión de entrenamiento (40 min). Los estudios previos han informado que VO2 responde más rápidamente a una transición escalonada desde el reposo a las intensidades submáximas después del entrenamiento (25), y que el aumento en VO2 es más rápido en los individuos entrenados que en los individuos no entrenados (26). En el presente estudio, el aumento en el VO2 en respuesta a la primera repetición de trabajo fue significativamente más largo que después de las repeticiones de trabajo subsiguientes. Esto coincide con los resultados de MacDonald et al. (20), quienes informaron que una sola repetición de ejercicios de alta intensidad mejoró el tiempo de aumento en el VO2 para la repetición subsiguiente de ejercicio (alta intensidad).
A pesar de las elevadas tasas de trabajo, hubo sólo una variación de 10 latidos.min-1 en la frecuencia cardíaca (HR) del ejercicio a lo largo de la sesión de entrenamiento y la recuperación de 60 s provocó una caída en la HR de 30-40 lat·min-1 (Figura 1B). La variación en la frecuencia cardíaca puede llegar a ser de 20 latidos-min-1 durante el ejercicio de alta intensidad de 20-60 min de duración en los individuos entrenados, a pesar de que las tasas de trabajo se mantienen iguales (14) y/o concentraciones plasmáticas de lactato estables o disminuidas (17). Esta limitación debe ser reconocida cuando se prescriben sesiones de entrenamiento de alta intensidad teniendo en cuenta solamente la HR (14).
No tenemos conocimiento de la existencia de algún otro estudio anterior que haya determinado aumentos en la temperatura muscular después de ejercicios de alta intensidad. Febbraio et al. (8) informaron previamente un aumento de 3,7°C en la temperatura muscular en varones altamente entrenados después de sólo 40 min de ciclismo de baja intensidad (225 W). Otra medida del control metabólico exhibido por nuestros ciclistas altamente entrenados durante la sesión de entrenamiento fue el menor incremento en la temperatura del músculo (3,2°C), después de la realización de un ejercicio con la misma duración que el ejercicio empleado en el estudio de Febbraio et al. (8), pero con tasas de trabajo relativa y absoluta notablemente superiores (es decir, 340 W, 85% VO2max). Es probable que, uno de los posibles mecanismos que producen tal observación sea un mayor flujo de sangre en el músculo durante los ejercicios de ciclismo de alta intensidad en comparación con el ciclismo de baja intensidad en nuestros sujetos altamente entrenados (7).
En conclusión, éste es el primer estudio que analizó las demandas metabólicas asociadas con una sesión de ejercicio intervalado aeróbico de alta intensidad en ciclistas de competición. Nuestros resultados revelan que estos atletas son capaces de mantener elevadas producciones de potencia aeróbica en estado estable, las cuales están asociadas con tasas altas de glucógenolisis y gasto de energía total. Las necesidades absolutas de combustible y las respuestas fisiológicas asociadas con este tipo de sesiones de entrenamiento intervalado son similares a las observadas cuando los ciclistas entrenados compiten en eventos que duran 60 min (13). Por lo tanto, no es sorprendente que tal entrenamiento se haya transformado en un elemento básico en el acondicionamiento para el ciclismo y haya sido asociado con mejoras en el rendimiento
Agradecimientos
Nos gustaría agradecer a María J. Brosnan y Angela Moquin por la ayuda técnica brindada durante las pruebas experimentales, y al Dr Louise Burke por la preparación de las dietas de los sujetos. Nosotros agradecemos a los sujetos y a sus entrenadores sobre todo por su excelente cooperación en esta investigación.
Referencias
1. ACEVEDO, E. O., and A. H. GOLDFARB (1989). Increased training intensity effects on plasma lactate, ventilatory threshold, and endurance. Med. Sci. Sports Exerc. 21:563568
2. BROOKS, G. A (1997). Importance of the crossover concept in exercise metabolism. Clin. Exp. Pharmacol. Physiol. 24:889894
3. COYLE, E. F (1995). Integration of physiological factors determining endurance performance ability. In: Exercise and Sport Science Reviews, Vol. 23, J. O. Holloszy (Ed.). Baltimore. Williams & Wilkins, pp. 2563
4. DILL, D. B., and D. L. COSTILL (1974). Calculation of percentage changes in volumes of blood, plasma and red cells in dehydration. J. Appl. Physiol. 37:247248
5. DYCK, D. J., C. T. PUTMAN, G. J. F. HEIGENHAUSER, E. HULTMAN, and L. L. SPRIET (1993). Regulation of fat-carbohydrate interaction in skeletal muscle during intense aerobic cycling. Am. J. Physiol. 265:E852E859
6. FEBBRAIO, M. A., M. F. CAREY, R. J. SNOW, C. G. STATHIS, and M. HARGREAVES (1996). Influence of elevated muscle temperature on metabolism during intense, dynamic exercise. Am. J. Physiol. 271: R1251R1255
7. FEBBRAIO, M. A., R. J. SNOW, C. G. STATHIS, M. HARGREAVES, and M. F. CAREY (1996). Blunting the rise in body temperature reduces muscle glycogenolysis during exercise in humans. Exp. Physiol. 81:685693
8. GORE, C. J., P. G. CATCHESIDE, S. N. FRENCH, J. M. BENNETT, and J. LAFORGIA (1997). Automated VO2max calibrator for open circuit indirect calorimetry systems. Med. Sci. Sports Exerc. 29:1095 1103
9. HAWLEY, J .A., L. M. BURKE, D. J. ANGUS, et al (1999). Effect of altering fat availability on metabolism and performance during intense exercise. Med. Sci. Sports Exerc. 31(5):S340
10. HAWLEY, J. A., K. H. MYBURGH, T. D. NOAKES, and S. C. DENNIS (1997). Training techniques to improve fatigue resistance and enhanced endurance performance. J. Sports Sci. 15:325333
11. HAWLEY, J. A., and T. D. NOAKES (1992). Peak power output predicts maximal oxygen uptake and performance time in trained cyclists. Eur. J. Appl. Physiol. 65:7983
12. HAWLEY, J. A., G. S. PALMER, and T. D. NOAKES (1997). Effects of 3 days of carbohydrate supplementation on muscle glycogen content and utilisation during 1-h cycling performance. Eur. J. Appl. Physiol. 75:407 412
13. JEUKENDRUP, A. E., and A. van DIEMEN (1998). Heart rate monitoring during training and competition in cyclists. J. Sports Sci. 16:S91 S99
14. JORDFELDT, L., A. JUHLIN-DANNFELT, and J. KARLSSON (1978). Lactate release in relation to tissue lactate in human skeletal muscle during exercise. J. Appl. Physiol. 44:350 352
15. KATCH, V. L., and F. I. KATCH (1974). A simple anthropometric method for calculating segmental leg limb volume. Res. Q. 45:211214
16. KINDERMAN, W., G. SIMON, and J. KEUL (1979). The significance of the aerobic-anaerobic transition for the determination of work load intensities during endurance training. Eur. J. Sports Med. 42:25 34
17. LINDSAY, F. H., J. A. HAWLEY, K. H. MYBURGH, H. H. SCHOMER, T. D. NOAKES, and S. C. DENNIS (1996). Improved athletic performance in highly trained cyclists after interval training. Med. Sci. Sports Exerc. 28:14271434
18. LOWRY, O. H., and J. V. PASSONNEAU (1974). A flexible system of enzymatic analysis. New York: Academic Press, Inc., pp. 199201
19. MACDONALD, M., P. K. PEDERSEN, and R. L. HUGHSON (1997). Acceleration of VO2 kinetics in heavy submaximal exercise by hyperoxia and prior high-intensity exercise. J. Appl. Physiol. 83:1318 1325
20. MARTIN, D., S. POWERS, M. CICALE, N. COLLOP, D. HUANG, and D. CRISWELL (1992). Validity of pulse oximetry during exercise in elite endurance athletes. J. Appl. Physiol. 72:455 458
21. NEVILL, M. E., G. C. BOGDANIS, L. H. BOOBIS, H. K.A. LAKOMY, and C. WILLIAMS (1996). Muscle metabolism and performance during sprinting. In: Biochemistry of Exercise IX, R. J. Maughan and S. M. Shirreffs (Eds.). Champaign, IL: Human Kinetics, pp. 243259
22. PASSONEAU, J. V., and V. R. LAUDERDALE (1974). A comparison of three methods of glycogen measurement in tissues. Anal. Biochem. 60:405 412
23. PERONNET, F., and D. MASSICOTTE (1991). Table of nonprotein respiratory quotient: an update. Can. J. Sports. Sci. 16:2329,
24. PHILLIPS, S. M., H. J. GREEN, M. J. MACDONALD, and R. L. HUGHSON (1995). Progressive effect of endurance training on VO2 kinetics at the onset of submaximal exercise. J. Appl. Physiol. 79:1914 1920
25. POWERS, S. K., S. DODD, and R. E. BEADLE (1985). Oxygen uptake kinetics in trained athletes differing in VO2max. Eur. J. Appl. Physiol. 54:306 308
26. POWERS, S. K., S. DODD, J. LAWLER, et al (1988). Incidence of respiratory induced hypoxemia in elite endurance athletes at sea level. Eur. J. Appl. Physiol. 58:289 302
27. ROMIJN, J. A., E. F. COYLE, J. HIBBERT, and R. R. WOLFE (1992). Comparison of indirect calorimetry and a new breath 13C/12C ratio, method during strenuous exercise. Am. J. Physiol. 263:E64E71
28. ROMIJN, J. A., E. F. COYLE, L. S. SIDOSSIS, et al (1993). Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. 28:E380E391
29. ROMIJN, J. A., E. F. COYLE, L. S. SIDOSSIS, X.-J. ZHANG, and R. R. WOLFE (1995). Regulation between fatty acid delivery and fatty acid oxidation during strenuous exercise. J. Appl. Physiol. 79:1939 1945
30. SAHLIN, K (1978). Intracellular pH and energy metabolism in skeletal muscle of man: with special reference to exercise. Acta Physiol. Scand. Suppl. 455:156
31. STEPTO, N. K., J. A. HAWLEY, S. C. DENNIS, and W. G. HOPKINS (1999). Effects of different interval-training programs on cycling timetrial performance. Med. Sci. Sports. Exerc. 31:736 741
32. WESTGARTH-TAYLOR, C., J. A. HAWLEY, S. RICKARD, K. H. MYBURGH, T. D. NOAKES, and S. C. DENNIS (1997). Metabolic and performance adaptations to interval training in endurance-trained cyclists. Eur. J. Appl. Physiol. 75:298 304
33. WESTON, A. R., K. H. MYBURGH, F. H. LINDSAY, S. C. DENNIS, T. D. NOAKES, and J. A. HAWLEY (1997). Skeletal muscle buffering capacity and endurance performance after high-intensity interval training by well-trained cyclists. Eur. J. Appl. Physiol. 75:713
34. WITHERS, R. T., N. P. CRAIG, P. C. BOURDON, and K. I. NORTON (1987). Relative body fat and anthropometric prediction of body density of male athletes. Eur. J. Appl. Physiol. 56:191200
35. COYLE, E. F., A. R. COGGAN, M. K. HOPPER, and T. J. WALTERS (1988). Determinants of endurance in well-trained cyclists. J. Appl. Physiol. 64:26222630
Cita Original
Stepto, N. K., D. T. Martin, K. E. Fallón, and J. A. Hawley. Metabolic demands of intense aerobic interval training in competitive cyclists. Med. Sci. Sports Exerc, Vol. 33, No. 2, 2001, pp. 303-310
Cita en PubliCE
John A Hawley, David T Martin, Nigel K Stepto Keiran E Fallon (2001). Demandas Metabólicas del Entrenamiento Intervalado Aeróbico de Alta Intensidad en Ciclistas de Competición. .https://g-se.com/demandas-metabolicas-del-entrenamiento-intervalado-aerobico-de-alta-intensidad-en-ciclistas-de-competicion-1360-sa-457cfb27202d73