
Cambios en el Tiempo de Activación de la Vía IGF-1/Akt/mTOR/p70s6k en el Músculo Gastrocnemio de Ratas Durante Series Repetidas de Ejercicio Excéntrico
Eisuke Ochi1, Naokata Ishii2, Koichi Nakazato31 Laboratory of Health and Sports Sciences, Center for Liberal Arts, Meiji Gakuin University.
2 Department of Life Sciences, Graduate School of Arts and Sciences, The University of Tokyo.
3 Department of Exercise Physiology, Graduate School of Sport Science, Nippon Sport Science University, Japan.
Resumen
El objetivo de este estudio fue examinar si los factores de crecimiento tipo insulínico 1 (IGF-1) y la actividad de la vía Akt/mTOR/p70S6K se ven alteradas por el ejercicio excéntrico crónico en el músculo gastrocnemio de ratas. Un grupo de ratas machos Wistar (n = 24) fue asignado de manera aleatoria a 1 de 2 grupos: grupo de ejercicio excéntrico (ECC) o grupo de ensayo controlado con intervención simulada (CON). A las ratas del grupo de ECC se las entrenó cada dos días durante 10 días (5 sesiones en total) o 20 días (10 sesiones en total). Después de 5 o 10 sesiones de ejercicios, se disecaron y pesaron los especímenes de músculo. La expresión de ARNm de IGF-1 y su variante, el factor de crecimiento mecánico sensible a la sobrecarga (MGF), se evaluó utilizando la reacción en cadena de la polimerasa en transcriptasa inversa (RT-PCR) en tiempo real. Las concentraciones tisulares de Akt (P), mTOR (P) y p70S6K (P) fueron medidas utilizando el análisis Western blot. No se observaron diferencias significativas en la masa muscular del gastrocnemio medial del grupo de ECC después de 5 sesiones de ejercicio, mientras que la masa muscular se incrementó significativamente después de 10 sesiones de ejercicio, con un incremento concomitante en el área de sección cruzada de las fibras musculares (p<0.05). La expresión de ARNm de IGF-1 y las concentraciones tisulares de Akt (P) y p70S6K (P) después de 10 sesiones de ejercicio fueron significativamente mayores que las de los controles combinados por edad y de las ratas que realizaron 5 sesiones de ejercicio. La expresión de ARNm de MGF en los grupos ECC5S y ECC10S fue significativamente mayor que la de los grupos de control en relación al mismo período de entrenamiento (p<0.01). Las concentraciones tisulares de mTOR (P) después de 10 sesiones de entrenamiento mostraron un incremento significativo en comparación con los controles en relación al mismo período de entrenamiento (p<0.01). Estos resultados sugieren que la activación de la vía de señalización IGF-1/Akt/mTOR/p70S6K se vuelve dominante en la fase posterior del ejercicio crónico, cuando se observa una hipertrofia muscular significativa.
Palabras Clave: vía de señalización, músculo esquelético, concentraciones excéntricas, hipertrofia
INTRODUCCION
Estudios previos han demostrado que el ejercicio incrementa la masa y la fuerza muscular (Armstrong et al., 1983; Wirth et al., 2003). La hipertrofia del músculo esquelético se debe al incremento del área de sección cruzada (hipertrofia) y a la cantidad de fibras musculares (hiperplasia). La hiperplasia y la hipertrofia están muy relacionadas con la proliferación/diferenciación de las células satélite y el incremento de la síntesis proteica (Appell et al., 1988; Giddings y Gonyea, 1992; Perry y Rudnick, 2000; Vierck et al., 2000).
La síntesis proteica en el músculo esquelético es promovida por los factores de crecimiento tipo insulínico 1 (IGF-1) (Barton-Davis et al., 1999). La activación de la vía de transducción de señales Akt/mTOR/p70S6K en parte es inducida por la respuesta autocrina/paracrina mediada por IGF-1, derivando en la hipertrofia muscular (Bodine et al., 2001; Glass 2003; Rommel et al., 2001). Los estímulos mecánicos, como el ejercicio, incrementan la expresión de IGF-1 y activan la vía de transducción de señales Akt/mTOR/p70S6K en gran medida (Burry et al., 2007; Sakamoto et al., 2002, Parkington et al., 2003; Spangenburg y McBride, 2006). Sakamoto et al. (2002) reportaron un incremento en la concentración de P-Akt luego de una hora de la realización de 1 serie de ejercicio en cinta ergométrica; mientras que la concentración de IGF-1 se incremento inmediatamente después de la realización de ejercicio con electro-estimulación (Sakamoto et al., 2002). Además, la fosforilación de la mTOR y la p70S6K, y el flujo descendente de Akt, se producen después de las contracciones musculares durante una única serie de ejercicio (Burry et al., 2007; Parkington et al., 2003; Spangenburg y McBride, 2006). Por sobre todo, una única serie de ejercicio induce a la síntesis proteica en el músculo ejercitado.
También se han analizado los efectos de la sobrecarga funcional crónica y del ejercicio sobre los cambios en la concentración de IGF-1 y la vía de transducción de señales Akt/mTOR/p70S6K (Adams y Haddad, 1996; Bodine et al., 2001; DeVol et al., 1990; Thomson et al., 2006). Con respecto a la sobrecarga funcional, la concentración de IGF-1 se incremento hasta transcurridos 7 días, pero disminuyó de allí en adelante (Adams y Haddad, 1996; DeVol et al.; 1990). Por otro lado, la activación de la vía Akt/mTOR/p70S6K aumentó de manera gradual después de más de una semana, y la activación de estas proteínas, en especial la p70S6K, concordó con la proporción de hipertrofia muscular (Bodine et al., 2001; Thomson et al., 2006). En conjunto, los niveles elevados de IGF-1 y la activación de Akt/mTOR/p70S6K parecieron estar correlacionados dentro del término de una semana posterior a una tenectomía. Pero luego de una semana, la activación de Akt/mTOR/p70S6K fue independiente a la IGF-1 en el modelo de sobrecarga funcional.
Con un modelo de ejercicio crónico, Hameed et al. (2004) demostraron que durante el ejercicio crónico las concentraciones de IGF-1 y sus variantes se elevaron después de 5 y 12 semanas de entrenamiento con sobrecarga, respectivamente, en personas ancianas (Hameed et al., 2004). Leger et al. (2006) reportaron que los niveles de Akt y mTOR fosforiladas se elevaron luego de 8 semanas de ejercicio muscular anaeróbico en seres humanos. En un modelo de ejercicios con animales, 20 días de ejercicio indujeron a una hipertrofia significativa, independientemente de la modalidad de contracción utilizada, i.e., excéntrica, isométrica o concéntrica (Adams et al., 2004). Por el contrario, las modalidades de ejercicio isométrico y concéntrico, pero no excéntrico, incrementaron la concentración muscular de IGF-1 y su variante, el factor de crecimiento mecánico sensible a la sobrecarga (MGF) (Adams et al., 2004). Si bien estos resultados sugieren que el ejercicio crónico induce una hipertrofia significativa, la elevación de los niveles de IGF-1 y la activación de la síntesis proteica, no se ha realizado una comparación directa de los cambios en los niveles de IGF-1, Akt, mTOR y p70S6K durante la misma modalidad de ejercicio crónico.
Se ha planteado la hipótesis de que el incremento de la masa muscular está asociado con la elevación del contenido de ARNm de IGF-1 y la activación de la vía Akt/mTOR/p70S6K. Previamente se han reportado incrementos significativos en la masa del músculo esquelético y en la fuerza muscular utilizando una modalidad de ejercicio excéntrico en el músculo gastrocnemio de ratas (Ochi et al., 2007). Por lo tanto, en este estudio, se utilizó este modelo para inducir un ejercicio excéntrico durante 10 y 20 días en el gastrocnemio medial de los sujetos. Después del ejercicio crónico, se midió la masa muscular húmeda, el área de sección cruzada de la fibra muscular (CSA), la expresión de ARNm de IGF-1 y la activación de la vía de transducción de señales Akt/mTOR/p70S6K.
METODOS
Animales
Un grupo de ratas machos Wistar (edad, 12 semanas; masa corporal, 303–365 g; n = 24) fue asignado aleatoriamente a 4 grupos: un grupo que realizó 5 sesiones de ejercicio excéntrico (ECC5S, n = 6); un grupo de control que realizó 5 sesiones de intervención simulada (CON5S, n = 6); un grupo experimental que realizó 10 sesiones de ejercicio excéntrico (ECC10S, n = 6); y un grupo de control que realizó 10 sesiones de intervención simulada (CON10S, n = 6). Los sujetos fueron ubicados de manera individual en una caja de acero a 22– 24°C y un ciclo de luz/oscuridad de 12-h. Se rasuró la pata derecha trasera de cada animal y durante el ejercicio se anestesió a los animales con isoflurano. El agua y la comida se encontraban disponibles ad libitum. No se hallaron diferencias significativas en la edad ni en la masa entre los grupos (como se muestra en la Tabla 1). El estudio fue aprobado por el Comité de Ética para la Experimentación Animal de la Universidad de Tokio.
CSA de las Fibras del Músculo Gastrocnemio Medial
Una parte de cada muestra congelada fue sujeto al análisis de tinción histoquímica con hematoxilina-eosina (H&E) (Sakura Finetek Japan Co. Japón). Previamente a la tinción, la muestra de músculo fue seccionada a un grosor de 10 µm a -20°C. Las secciones de cada grupo se colocaron en el mismo portaobjetos de vidrio a fin de eliminar la variabilidad en las condiciones de tinción. Posteriormente a la tinción, se observó a las fibras bajo la luz de un microscopio. El CSA de las fibras musculares se determinó utilizando una imagen del NIH (versión 1.61; National Institutes of Health, EUA). Se midieron 200 fibras de cada animal.
Ejercicio Excéntrico para el Músculo Gastrocnemio Medial
El sistema de ejercicios excéntricos para la articulación del tobillo de las ratas fue similar al sistema descrito en el estudio previo (Ochi et al., 2007). Los animales fueron anestesiados y se los sujetó con firmeza a la plataforma de un dinamómetro isocinético en posición de decúbito prono, y se estimuló eléctricamente el músculo tríceps sural de la pata derecha trasera. A las ratas del grupo que realizó ejercicio excéntrico se las entrenó cada dos días durante 10 días (5 sesiones en total; ECC5S) o 20 días (10 sesiones en total; ECC10S). El voltaje de estimulación se ajustó para producir un torque de contracción isométrica submáximo a través de un electrodo adhesivo de superficie (7.5 mm × 7.5 mm) conectado a un estimulador y aislante eléctrico (Nihon Koden, Japón). Luego se estimuló al músculo durante 3 s para provocar una contracción tetánica submáxima. Un segundo después del inicio de la estimulación, se le realizó a la articulación del tobillo una dorsiflexión isocinética forzada para provocar una contracción excéntrica del músculo tríceps sural. La velocidad y el rango de estiramiento forzado fueron de 30°/s y de 0° (definido como el ángulo en el cual la planta del pie y el hueso tibial se ubican ortogonalmente) a 45°, respectivamente. Cada sesión de ejercicio consistió de 4 series de 5 contracciones, con un intervalo de 5-min entre cada serie. A los grupos que realizaron la intervención simulada (CON5S y CON10S) sólo se los anestesió pero no se los ejercitó. Dentro de las 12 hs posteriores a la última sesión de ejercicios, los especímenes de músculo se disecaron, se pesaron y de inmediato se congelaron en N2 líquido y se almacenaron a -80°C hasta realizar el análisis.
Análisis de ARNm por medio de la Reacción en Cadena de la Polimerasa en Transcriptasa Inversa (RT-PCR) en Tiempo Real
Las expresiones de ARNm de IGF-1 y MGF en el músculo gastrocnemio medial se midieron utilizando (RT-PCR) en tiempo real, tal como lo describieran Nakazato et al. (2007). El aislamiento del ARN y los métodos de RT-PCR se realizaron según las instrucciones del fabricante (RNeasy MiniKit, QIAGEN, EUA, High-capacity cDNA Archive kit, Applied Biosystems, EUA); la RT-PCR se llevó a cabo con una mezcla maestra en un volumen final de 12.5 µl (SYBR Green PCR Master Mix, Applied Biosystems, EUA). Se realizó una amplificación en un sistema ABI Prism (Applied Biosystems, EUA) utilizando el siguiente programa: 2 min a 50°C y 10 min a 95°C, seguidos de 15 s a 95°C y 1 min a 60°C para 40 ciclos. Las curvas externas estándar se utilizaron para un análisis cuantitativo, y los niveles de ARNm de cada gen objetivo se normalizaron a los de 18 s, los cuales sirvieron de control. Los cebadores utilizados fueron 5′-GCATTGTGGATGAGTGTTGC-3′ (cebador directo) y 3′-GGCTCCTCCTACATTCTGTA-5′ (cebador inverso) para IGF-1 (GenBank: X06043) y 5′-GCATTGT GGATGAGTGTTGC-3′ (cebador directo) y 3′-CTTTTCTTGTGTGTCGATAGG-5′ (cebador inverso) para MGF-1 (GenBank: X06108) en base a Adams et al. (2004). Dado que el amplicon varía en la región de intercambio de las variables de IGF, se analizaron las mezclas de todos los ARNm de las variables de IGF por medio de este conjunto de cebadores. Los datos se analizaron utilizando un método comparativo de umbral crítico (Ct) donde la cantidad de genes objetivo se normalizó a la del gen control endógeno.
Análisis de los Indicadores de Transducción de Señales por medio de Western Blotting
Las muestras del músculo gastrocnemio medial se maceraron en N2 líquido y se homogeneizaron en mM Tris-Cl (pH 7.4), 150 mM NaCl, 5 mM EDTA, 0.5% dodecil sulfato de sodio (SDS), 1% desoxicolato, 0.1% tritón X-100, 1% Nonidet P-40 (NP-40), 0.05% mercaptoetanol, 10 mg/mL PMSF, 0.5 mg/mL leupeptina, 0.2 mg/mL aprotinina, y 1mM Na3VO4.
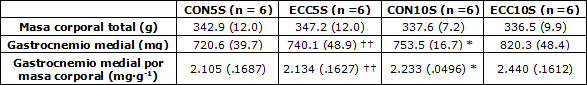
Tabla 1. Masa corporal total, masa muscular húmeda y masa muscular húmeda relativa a la masa corporal. Los datos son medias (± DE). *p < 0.05 CON10S vs. ECC10S, †† p< 0.01 ECC5S vs. ECC10S.
El homogenado se centrífugo a 15,000 × g durante 25 min a 4°C. Las concentraciones proteicas se determinaron utilizando un kit para la determinación de la concentración proteica (Protein Assay II, Bio-Rad, Richmond, CA, EUA). Se mezcló un equivalente de 25 µg del extracto total de proteína de cada muestra con el amortiguador de la muestra, se hirvió y se cargó en geles de poliacrilamida-SDS. Las proteínas se separaron por medio de PAGE discontinuo a través de geles de acrilamida al 12.5%. Las muestras se separaron electroforeticamente a 180mA durante 90 min y luego se transfirieron a membranas PVDF (ATTO, Japan). Las membranas se bloquearon con PBS que contenía 5% de leche descremada y se incubaron durante 1h. Todos los anticuerpos primarios se incubaron durante la noche a 4°C: un anticuerpo monoclonal anti-Akt (P) (Cell Signaling Technology, EUA), un anticuerpo policlonal anti-mTOR (P) (Cell Signaling Technology, EUA) y un anticuerpo monoclonal anti-p70S6K (Cell Signaling Technology, EUA), cada uno diluido a 1/1,000. Las membranas se lavaron e incubaron durante 10 min 2 veces a temperatura ambiente. Se utilizó un HRP-conjugado IgG de cabra anti-ratón o anti-conejo secundario en una disolución de 1:10,000 (Santa-Cruz, Biotechnology, EUA). La actividad HRP se detectó mediante una quimioluminiscencia (Supersignal West Dura; Pierce, EUA) utilizando un detector de quimioluminiscencia (AE6961, ATTO, Japón) y se cuantificó utilizando un analizador de CS (ATTO). A fin de reducir al máximo el efecto de las condiciones de reacción, las muestras de cada gel contenían cada uno de los grupos experimentales.
Análisis Estadísticos
Todos los valores están expresados como medias y desviaciones estándar. Se utilizó el análisis de varianza (ANOVA) de una vía a fin de comparar el CSA de las fibras musculares en 10 sesiones. Se utilizó un ANOVA de dos vías seguido de un test de Bonferroni para el análisis de la masa corporal total, de la masa muscular húmeda, de la masa muscular húmeda por masa corporal, de la expresión de ARNm de IGF-1 y de las concentraciones tisulares de Akt (P), mTOR (P) y p70S6K (P). El nivel de significancia se estableció en p<0.05.
RESULTADOS
La Masa Húmeda y la Masa Húmeda Relativa de los Músculos y CSA de las Fibras Musculares
Ni la masa muscular del gastrocnemio medial ni la masa muscular por masa corporal en el grupo ECC5S se incrementaron de manera significativa, mientras que las del grupo ECC10S fueron significativamente mayores que las del grupo CON10S, con un incremento concomitante en el CSA de las fibras musculares (p<0.05; Tabla 1 y Figura 1). Además, los valores del grupo ECC10S también fueron significativamente mayores que los del grupo ECC5S (p<0.01).
Expresión de ARNm de IGF-1 y ARNm de MGF
La expresión de ARNm de IGF-1 en el gastrocnemio se determinó mediante RT-PCR, y se halló que fue significativamente mayor en el grupo ECC10S que en los grupos CON10S y ECC5S (p<0.05); (Figura 2A). Por otro lado, las expresiones de ARNm de MGF en los grupos ECC5S y ECC10S fueron significativamente mayores que las de cada grupo control cuando se compararon los mismos períodos de entrenamiento (p < 0.01).
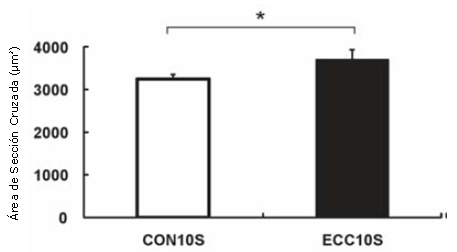
Figura 1. Área de sección cruzada de las fibras musculares (CSA) en el músculo gastrocnemio medial determinada a través de tinción H&E. Los valores son medias ± DE. Cantidad total de fibras = 1200 para cada grupo. * p<0.05.
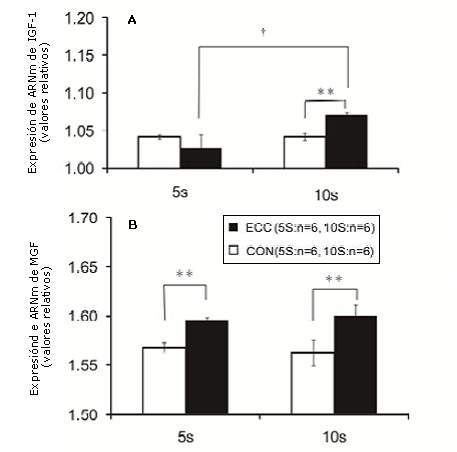
Figura 2. Expresión de ARNm muscular para IGF-1 (A) o MGF( B) determinada mediante la utilización de RT-PCR en tiempo real. La expresión de ARNm de IGF-1 en el grupo ECC10S fue significativamente mayor que la de CON10S y ECC5S. La expresión de ARNm de MGF en los grupos ECC5S y ECC10S fue significativamente mayor que la de cada grupo control en relación al mismo período de entrenamiento (CON5S o CON10S, respectivamente). Los valores son medias ± DE. * p<0.05, ** p<0.01, grupos de entrenamiento excéntrico versus los grupos de intervención simulada. † p<0.05, ECC10S vs ECC5S.
Fosforilación de las Proteínas de Transducción de Señales
Las concentraciones de tejido de Akt (P), mTOR (P) y p70S6K (P) se determinaron utilizando el análisis por Western Blot. La concentración tisular de Akt (P) en el grupo ECC10S fue significativamente más elevada que la de los grupos CON10S y ECC5S (p<0.01; Figura 3). La concentración tisular de mTOR (P) en el grupo ECC10S exhibió un incremento significativo en comparación con controles de la misma edad (p<0.01; Figura 4). La concentración tisular de p70S6K en el grupo ECC10S fue significativamente más elevada que la observada en los grupos CON10S y ECC5S (p<0.001; Figura 5).
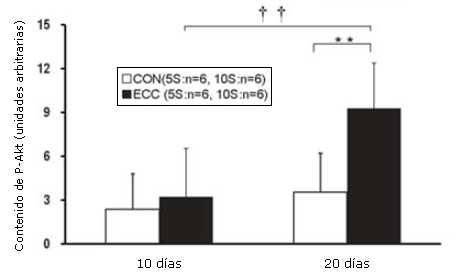
Figura 3. Contenido muscular de la Akt(P) determinado por medio Western Blotting. Los valores son medias ± DE. **p<0.01, ECC10S vs CON10S. †† p<0.01, ECC10S vs ECC5S.
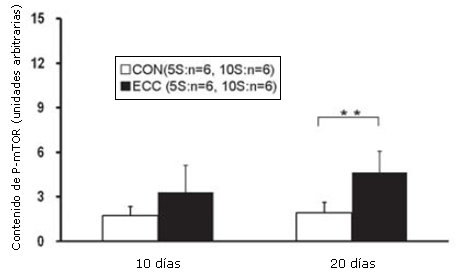
Figura 4. Contenido muscular de la mTOR(P) determinado por medio de Western Blotting. Los valores son medias ± DE. ** p<0.01, ECC10S vs CON10S.
DISCUSION
Mediante la utilización de este modelo de ejercicio, se ha demostrado que 20 días, pero no 10 días, de ejercicio excéntrico produce un incremento en la masa muscular (Tabla 1). Dado que no se han observado cambios en la masa muscular del gastrocnemio medial ni en el índice de músculo por masa corporal después de 5 sesiones de ejercicio, se concluye que 5 sesiones de ejercicio son inadecuadas para provocar la hipertrofia muscular con este modelo. Por otro lado, se observó una hipertrofia significativa después de 10 sesiones de ejercicio, y estuvo acompañada por un incremento significativo en la expresión de ARNm de IGF-1 y la activación de la vía Akt/mTOR/p70S6K. En conjunto, estos resultados sugieren que la vía de señalización IGF-1/Akt/mTOR está asociada a una significativa hipertrofia muscular en el ejercicio crónico. Los futuros estudios que utilicen este modelo deberían poner más atención en el tiempo de disección. Las disecciones de este estudio se realizaron dentro de las 12 hs posteriores a la última sesión de ejercicios. Los presentes resultados sugieren que los incrementos observados en la ARNm, la mTOR (P), la Akt (P) y la p70S6k (P) de la IGF-1 después de 10 sesiones de ejercicio pueden mantenerse por hasta 12 hs.
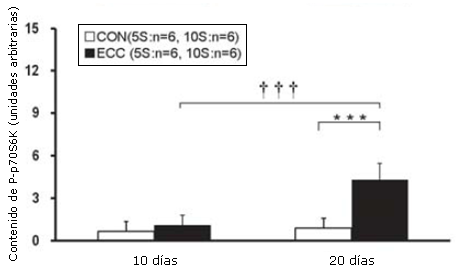
Figura 5. Contenido muscular de la p70S6K(P) determinado por medio de Western Blotting. Los valores son medias ± DE. *** p<0.001, ECC10S vs CON10S. ††† p<0.001, ECC10S vs ECC5S.
En este estudio, se cuantificó la expresión de ARNm de IGF-1. Aunque no se observaron cambios después de las 5 sesiones de ejercicio, se observó un aumento significativo después de 10 sesiones de ejercicio (p<0.05; Figura 2A). Estos resultados concuerdan con las respuestas de IGF-1 posteriores a la realización de ejercicios que provocan una respuesta anabólica en seres humanos (Bamman et al., 2001; Hameed et al., 2004). Las series repetidas de contracciones excéntricas del gastrocnemio en un modelo con ratas también indujeron una expresión de ARNm de IGF-1 más elevada después de 10 sesiones de ejercicio en comparación con lo observado en los controles, pero la diferencia que se observó no fue estadísticamente significativa (Adams et al., 2004). En el presente estudio, se observó una significativa expresión de ARNm de IGF-1 después de 10 sesiones de ejercicio excéntrico. Dado que el rango de estiramiento forzado utilizado en el presente modelo y el utilizado por Adams et al. (2004) fueron diferentes (45° y 20°, respectivamente), la expresión de MGF tanto después de 5 como de 10 sesiones mostraron significativos incrementos (Figura 2B). Éste no fue el caso del estudio de Adams et al (2004). Dado que el MGF se definió como la isoforma de la IGF-1 sensible a la carga, el incremento de MGF podría ser a causa de una mayor carga mecánica en el presente estudio y en comparación con Adams et al (2004).
El cambio en el transcurso de tiempo en la expresión de ARNm de IGF-1 es otro punto a considerar. En un modelo de sobrecarga funcional, la IGF-1 sólo se observó en el término de 1 semana (Adams y Haddad 1996; Ishido et al., 2008). Dado que el tiempo de incremento en la expresión de ARNm de IGF-1 se asoció con un incremento significativo en la hipertrofia muscular, se ha planteado la hipótesis de que la activación de la IGF-1 cumple un papel importante en el inicio de la hipertrofia muscular durante el ejercicio crónico de fuerza.
Estudios previos han demostrado que una única sesión de ejercicio muscular provoca un incremento transitorio en la activación de la vía de señalización Akt/mTOR/p70S6K dentro de los músculos que se han ejercitado (Burry et al., 2007; Parkington et al., 2003; Sakamoto et al., 2002; Spangenburg y McBride, 2006). Sin embargo, la relevancia de la relación entre la concentración de IGF-1 y la activación de la vía Akt/mTOR/p70S6K para la hipertrofia muscular es controversial. Estudios previos han analizado la relación entre IGF-1 y Akt/mTOR/p70S6K utilizando modelos de carga funcional y ejercicio (Adams 2002; Hornberger et al., 2006; Ishido et al., 2008; O’Neil et al., 2009; Spangenburg et al., 2008). Spangenburg et al (2008) reportaron que la activación del receptor de IGF-1 no fue necesario para inducir hipertrofia muscular y no tuvo influencia sobre la activación de la Akt y la p70S6k (Spangenburg et al., 2008). Además, se ha demostrado que la fosfolipasa D (PLD) y el ácido fosfatídico (PA) activan la mTOR y la p70S6K, independientemente de los IGF-1 (Hornberger et al., 2006). Por el contrario, se ha mostrado que los IGF-1 activan la vía Akt/mTOR/p70S6K en los modelos de ejercicio crónico (Adams 2002; Ishido et al., 2008). Aunque se cree que la IGF-1 y la vía Akt/mTOR/p70S6K cumplen un papel importante en la hipertrofia muscular, también se plantea la hipótesis de la presencia de vías redundantes tales como PLD y PA.
CONCLUSION
En el presente estudio se mostró que el modelo de ejercicio muscular provocó un incrementó la masa húmeda del músculo gastrocnemio medial y en el CSA de las fibras musculares. Estos incrementos sólo podrían lograrse después de 10 sesiones de ejercicios (20 días). La expresión incrementada de ARNm de IGF-1 estuvo acompañada de la fosforilación de la Akt, la mTOR y la p70S6K, y dichos eventos moleculares sólo se observaron luego de 10 sesiones de ejercicio. Estos resultados, junto con los de los estudios previos, sugieren que la activación vía IGF-1/Akt/mTOR/p70S6K cumple un papel importante en fases posteriores del ejercicio crónico, cuando se observa una hipertrofia muscular significativa.
Puntos Clave
- Se confirmó que el modelo de ejercicio muscular utilizado en ratas y que utiliza un equipo de desarrollo original incrementó la masa húmeda del músculo gastrocnemio medial y las áreas de sección cruzada de las fibras musculares en 10 sesiones (20 días), pero no en 5 sesiones (10 días).
- Se aclaró que los incrementos de masa muscular y el CSA de las fibras musculares estuvieron acompañados por la expresión de ARNm de IGF-1, y de la concentración de Akt, mTOR y p70S6K fosforiladas.
- Estos resultados sugieren que la hipertrofia muscular del presente modelo se logró después de 10 sesiones de ejercicio y se asoció a la activación de la vía de señalización IGF-1/Akt/mTOR/p70S6K.
AGRADECIMIENTOS
Se agradece a Tatsuro Hirose por el útil debate en la escritura del manuscrito. El presente estudio fue financiado por la Subvención para Científicos Jóvenes (B; 21700663) y para la Investigación Científica (B; 21300235, C; 00307993) de KAKENHI.
Referencias
1. Adams, G.R (2002). Invited review: Autocrine/paracrine IGF-I and skeletal muscle adaptation. Journal of Applied Physiology 93, 1159-1167
2. Adams, G.R., Cheng, D.C., Haddad, F. and Baldwin, K.M (2004). Skeletal muscle hypertrophy in response to isometric, lengthening, and shortening training bouts of equivalent duration. Jour-nal of Applied Physiology 96, 1613-1618
3. Adams, G.R. and Haddad, F (1996). The relationships among IGF-1, DNA content, and protein accumulation during skeletal muscle hypertrophy. Journal of Applied Physiology 81, 2509-2516
4. Appell, H.J., Forsberg, S. and Hollmann, W (1988). Satellite cell activation in human skeletal muscle after training: Evidence for muscle fiber neoformation. International Journal of Sports Medicine 9, 297-299
5. Armstrong, R.B., Ogilvie, R.W. and Schwane, J.A (1983). Eccentric exercise-induced injury to rat skeletal muscle. Journal of Ap-plied Physiology: Respiratory, Environmental and Exercise Physiology 54, 80-93
6. Bamman, M.M., Shipp, J.R., Jiang, J., Gower, B.A., Hunter, G.R., Goodman, A., McLafferty, J.L.Jr. and Urban, R.J (2001). Me-chanical load increases muscle IGF-I and androgen receptor ARNm concentrations in humans. American Journal of Physiology, Endocrinology and Metabolism 280, E383-390
7. Barton-Davis, E.R., Shoturma, D.I. and Sweeney, H.L (1999). Contribu-tion of satellite cells to IGF-I induced hypertrophy of skeletal muscle. Acta Physiologica Scandinavica 167, 301-305
8. Bodine, S.C., Stitt, T.N., Gonzalez, M., Kline, W.O., Stover, G.L., Bauerlein, R., Zlotchenko, E., Scrimgeour, A., Lawrence, J.C., Glass, D.J. and Yancopoulos, G.D (2001). Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nature Cell Biology 3, 1014-1019
9. Burry, M., Hawkins, D. and Spangenburg, E.E (2007). Lengthening contractions differentially affect p70s6k phosphorylation compared to isometric contractions in rat skeletal muscle. European Journal of Applied Physiology 100, 409-415
10. DeVol, D.L., Rotwein, P., Sadow, J.L., Novakofski, J. and Bechtel, P.J (1990). Activation of insulin-like growth factor gene expression during work-induced skeletal muscle growth. The American Journal of Physiology 259, E89-95
11. Giddings, C.J. and Gonyea, W.J (1992). Morphological observations supporting muscle fiber hyperplasia following weight-lifting exercise in cats. The Anatomical Record 233, 178-195
12. Glass, D.J (2003). Molecular mechanisms modulating muscle mass. Trends in Molecular Medicine 9, 344-350
13. Hameed, M., Lange, K.H., Andersen, J.L., Schjerling, P., Kjaer, M., Harridge, S.D. and Goldspink, G (2004). The effect of recombinant human growth hormone and resistance training on IGF-I ARNm expression in the muscles of elderly men. Journal of Physiology 555, 231-240
14. Hornberger, T.A., Chu, W.K., Mak, Y.W., Hsiung, J.W., Huang, S.A. and Chien, S (2006). The role of phospholipase D and phos-phatidic acid in the mechanical activation of mTOR signaling in skeletal muscle. Proceedings of the National Academy of Sci-ences of the United States of America 103, 4741-4746
15. Ishido, M., Kasuga, N. and Masuhara, M (2008). Time course changes of the expression of IGF-I, phosphorylated akt and phosphorylated mTOR in myofibers of the early stage of functionally overloaded skeletal muscle. Advances in Exercise and Sports Physiology 14, 25-29.
16. Leger, B., Cartoni, R., Praz, M., Lamon, S., Deriaz, O., Crettenand, A., Gobelet, C., Rohmer, P., Konzelmann, M., Luthi, F. and Rus-sell, A.P (2006). Akt signalling through GSK-3beta, mTOR and Foxo1 is involved in human skeletal muscle hypertrophy and at-rophy. The Journal of Physiology 576, 923-933
17. Nakazato, K., Song, H., Waga, T (2007). Dietary apple polyphenols enhance gastrocnemius function in wistar rats. Medicine and Science in Sports and Exercise 39, 934-940
18. Ochi, E., Nakazato, K. and Ishii, N (2007). Effects of eccentric exercise on joint stiffness and muscle connectin (titin) isoform in the rat hindlimb. The Journal of Physiological Sciences 57, 1-6
19. ONeil, T.K., Duffy, L.R., Frey, J.W. and Hornberger, T.A (2009). The role of phosphoinositide 3-kinase and phosphatidic acid in the regulation of mammalian target of rapamycin following eccentric contractions. The Journal of Physiology 587, 3691-3701
20. Parkington, J.D., Siebert, A.P., LeBrasseur, N.K. and Fielding, R.A (2003). Differential activation of mTOR signaling by contractile activity in skeletal muscle. American Journal of Physiol-ogy.Regulatory, Integrative and Comparative Physiology 285, R1086-1090
21. Perry, R.L. and Rudnick, M.A (2000). Molecular mechanisms regulating myogenic determination and differentiation. in Bio-science: A Journal and Virtual Library 5, D750-767
22. Rommel, C., Bodine, S.C., Clarke, B.A., Rossman, R., Nunez, L., Stitt, T.N., Yancopoulos, G.D. and Glass, D.J (2001). Mediation of IGF-1-induced skeletal myotube hypertrophy by PI(3)K/Akt/mTOR and PI(3)K/Akt/GSK3 pathways. Nature Cell Biology 3, 1009-1013
23. Sakamoto, K., Hirshman, M.F., Aschenbach, W.G. and Goodyear, L.J (2002). Contraction regulation of akt in rat skeletal muscle. The Journal of Biological Chemistry 277, 11910-11917
24. Spangenburg, E.E., Le Roith, D., Ward, C.W. and Bodine, S.C (2008). A functional insulin-like growth factor receptor is not necessary for load-induced skeletal muscle hypertrophy. The Journal of Physiology 586, 283-291
25. Spangenburg, E.E. and McBride, T.A (2006). Inhibition of stretch-activated channels during eccentric muscle contraction attenuates p70S6K activation. Journal of Applied Physiology 100, 129-135
26. Thomson, D.M. and Gordon, S.E (2006). Impaired overload-induced muscle growth is associated with diminished translational signalling in aged rat fast-twitch skeletal muscle. The Journal of Physiology 574, 291-305
27. Vierck, J., O'Reilly, B., Hossner, K., Antonio, J., Byrne, K., Bucci, L. and Dodson, M (2000). Satellite cell regulation following myotrauma caused by resistance exercise. Biology International 24, 263-272
28. Wirth, O., Gregory, E.W., Cutlip, R.G. and Miller, G.R (2003). Control and quantitation of voluntary weight-lifting performance of rats. Journal of Applied Physiology 95, 402-412
Cita Original
Eisuke Ochi, Naokata Ishii and Koichi Nakazato. Time Course Change of IGF-1/Akt/mTOR/p70s6K Pathway Activation in Rat Gastrocnemius Muscle during Repeated Bouts of Eccentric Exercise. Journal of Sports Science and Medicine (2010) 9, 170 - 175.
Cita en PubliCE
Eisuke Ochi, Naokata Ishii Koichi Nakazato (2010). Cambios en el Tiempo de Activación de la Vía IGF-1/Akt/mTOR/p70s6k en el Músculo Gastrocnemio de Ratas Durante Series Repetidas de Ejercicio Excéntrico. .https://g-se.com/cambios-en-el-tiempo-de-activacion-de-la-via-igf-1-akt-mtor-p70s6k-en-el-musculo-gastrocnemio-de-ratas-durante-series-repetidas-de-ejercicio-excentrico-1269-sa-i57cfb271e07ec