Bicentenario de las Investigaciones sobre el Lactato en el Músculo
Bruce L Gladden0 Department of Kinesiology, Auburn University, Auburn, AL, Estados Unidos.
Resumen
El año 2008, se cumplió el bicentenario de las investigaciones sobre el metabolismo muscular del lactato. Berzelius vinculó la acumulación de lactato con el ejercicio en 1807/1808 cuando observó la presencia de lactato en los músculos de "ciervos cazados" Hoy, el mecanismo exacto de la oxidación intramuscular de lactato y la relación de la lactato deshidrogenasa con las mitocondrias no han sido establecidos y forman parte de las discusiones sobre el transporte de lactato.
Palabras Clave: transporte intracelular de lactato, lactato deshidrogenasa, láctico deshidrogenasa, isoenzimas, antecedentes
INTRODUCCION
Descubrimiento del Lactato
El lactato o ácido láctico fue descubierto a partir de leche agria por un ayudante de farmacia sueco, Carl Wilhelm Scheele (1742-1786), en 1780 (2[pp6-8]). Benninga (2[pp7-8]) aportó una traducción moderna de la técnica de aislamiento de Scheele; que citamos parcialmente aquí: ' El suero agrio se evapora a un octavo de su volumen, posteriormente la cuajada [proteínas de leche precipitadas] se remueve por filtración. El filtrado se satura con leche de cal [hidróxido de calcio], se filtra nuevamente y se diluye con tres veces su volumen de agua' (2). Después de una serie de pasos adicionales, Scheele concluyó que (traducción moderna): ' El ácido láctico producido es tan puro como el que puede obtenerse mediante reacciones químicas ' (2). El nuevo ácido fue nombrado ' Mjölksyra' que significa “ácido de leche” (2[p8]). El lactato (La-) se encuentra como ingrediente ácido de productos lácteos agrios, frutas y verduras fermentadas y salchichas (2[p1]) y como un modificador o resaltador de sabor en las bebidas carbonatadas. También se ha utilizado en la curtiembre de cueros, en la tinción ácida de lanas y como producto farmacéutico en forma de lactato de calcio; sus ésteres han sido utilizados como solventes de laca y una pequeña cantidad ha sido utilizada en plásticos. El libro de Benninga (2) relata detalladamente la historia del (La-) y su producción a gran escala con propósitos industriales desde una perspectiva biotecnológica, lo que aporta un punto de vista que está completamente fuera del razonamiento de la mayoría de los científicos relacionados con la biología. Es importante destacar que en los niveles fisiológicos de pH en el músculo y en la sangre, más del 99% del ácido láctico se encuentra disociado en el anión (La-) y protones (H+); por consiguiente, en esta revisión lo denominaremos en general 'La-'.
Scheele también descubrió el cloro, manganeso, ácido arsénico y fluoruro de silicio junto con otros numerosos compuestos (28[p461]). Sin embargo, su mayor reconocimiento proviene de su trabajo con el O2 donde fue el primero en demostrar que el aire está compuesto de dos componentes y que uno de ellos produce la combustión (28[p461]). Scheele descubrió el O2 en 1772, más de un año antes que Joseph Priestley (1733-1804). Desgraciadamente, sus experimentos (A chemical treatise on air and fire; Tratado Químico sobre el aire y el fuego) no fue publicado hasta 1777, probablemente debido a la dilación de su amigo y defensor Torbern Bergman (1735-1784), quién escribió la introducción al libro (28[pp49,461]). Como resultado, él comparte el mérito del descubrimiento del O2 con Priestley.
LACTATO Y EJERCICIO
Aunque Scheele es recordado como el descubridor del La-, fue Jöns Jacob Berzelius (1779-1848), otro sueco, quien relacionó indeleblemente al (La-) con el estudio del ejercicio, cuando informó la presencia de La- en los músculos de ciervos cazados en 1807 (19[p41]) o 1808 (20[pxxvii]). Berzelius aparentemente se convenció de que la cantidad de La- en un músculo era proporcional a la cantidad de ejercicio que el músculo había realizado (19[p41]).
No se menciona frecuentemente que Berzelius también descubrió el ácido pirúvico (28[p56]). En ese sentido, es digno que nosotros hagamos una pausa en el bicentenario de la investigación sobre el metabolismo del lactato en el músculo, para realizar una apreciación global histórica breve y una discusión sobre las controversias actuales. En la actualidad existen por lo menos tres áreas de discusión sobre el estudio fisiológico /bioquímico del La-: 1) el papel del La- como un agente causal o de prevención de la fatiga muscular, 2) el papel de La- en la acidosis del ejercicio de alta intensidad y 3) la vía del metabolismo intracelular de La- en el músculo. Esta revisión se centrará en el tercer punto.
El metabolismo del La- en el músculo durante y después del ejercicio es de especial importancia dado que el músculo esquelético no sólo es la fuente más importante de La- si no que también es el principal consumidor de La-. Por ejemplo, durante la recuperación del ejercicio intenso, la oxidación es la vía principal para la remoción del La-, particularmente en el músculo esquelético.
LACTATO DESHIDROGENASA: DESCUBRIMIENTO E ISOENZIMAS
La discusión sobre el metabolismo intracelular del La- no sólo es una historia sobre el La- si no que también sobre la enzima que cataliza su formación, la enzima lactato deshidrogenasa (LDH). Berzelius tiene una relación lejana con este aspecto de la historia, porque él fue el primero (en 1836) en proponer el término catálisis, que él utilizaba para explicar la acción de compuestos que facilitaban las reacciones químicas sin experimentar algún cambio ellos mismos (19[pp30-31]). Luego sobrevino un largo período durante el cual los científicos estudiaron la capacidad para promover las reacciones que tenían 'los fermentos', los tejidos o extractos de tejidos de diferentes organismos. En ese momento, los fermentos fueron divididos en dos grupos: 1) aquellos que eran desorganizados o solubles y 2) aquéllos que eran organizados o insolubles (es decir, estaban compuestos por microorganismos intactos).
En 1877, Willy Kühne (1837-1900) propuso el nombre 'enzimas' (griego, 'en levaduras') para los fermentos desorganizados (28[pp300-301]). Veinte años después, Eduard Buchner (1860-1917) molió células de levadura con arena a una temperatura controlada y obtuvo un extracto que estaba libre de células de levadura pero todavía conservaba la capacidad de fermentar sacarosa a etanol, demostrando concluyentemente que las células no eran esenciales para la fermentación (28[pp87-88]).
Posteriormente en 1926, el americano James Batcheller Sumner (1877-1955) aisló por primera vez la forma cristalina de una enzima, la ureasa, a partir de porotos (28[p498-499]).
Quizás no sorprendentemente, fue Otto Meyerhof (1884-1951) (28[p368]) en 1919, estudiando el músculo esquelético, quien demostró por primera vez la acción de la LDH. Después de quizás cientos de investigaciones en el tema de LDH, aisló LDH pura en forma cristalina a partir de corazones de bueyes.
En el comienzo de los años cincuenta, varios investigadores aportaron evidencia que las preparaciones de LDH eran heterogéneas. Este trabajo culminó en 1959 cuando Markert y Møller (17), usando el método de la separación superior de electroforesis en gel de almidón junto con técnicas de tinción histoquimicas, demostraron claramente cinco formas diferentes de la enzima LDH. Ellos (17) primero también propusieron el término isoenzima para designar a las diferentes formas moleculares de una enzima. En 1962, el grupo de Kaplan (6) propuso la hipótesis del modelo de organización tetramérica de las isoenzimas de LDH que todavía se aplica hoy: existen dos sub-unidades monoméricas, H (corazón) y M (músculo) que puede combinarse en grupos de cuatro para producir cinco especies moleculares diferentes; HHHH, HHHM, HHMM, HMMM y MMMM. La nomenclatura moderna denomina a los monómeros como A (músculo) y B (corazón) lo que da origen a LDH1 (B4), LDH2 (B3A1), LDH3 (B2A2), LDH4 (B1A3) y LDH5 (A4). El laboratorio de Kaplan (6) avanzó para proponer que las diferentes propiedades de las isoenzimas eran fisiológicamente importantes en la regulación del metabolismo celular. Específicamente, propusieron que la sensibilidad de las formas relacionadas al corazón (H4, H3M1), frente a la inhibición del piruvato direccionaba al piruvato hacia una oxidación en un ambiente aeróbico como el corazón, mientras que la relativa insensibilidad a la inhibición del piruvato, de las formas asociadas al músculo (H1M3, M4) facilitaba la degradación anaeróbica de los carbohidratos a lactato en los músculos que realizaban ejercicios (6).
Este esquema ha sido apoyado por evidencia circunstancial. Por ejemplo, la capacidad del músculo para utilizar La- se correlaciona, tanto con la capacidad oxidativa como con el contenido de LDH del tipo del corazón. Como se observa en la revisión de Van Hall (27), también hay evidencia que indica que las fibras oxidativas de los músculos tienen aproximadamente la mitad de la actividad total de LDH de las fibras glucolíticas y de esa actividad, una cantidad relativa mayor la tienen las formas relacionadas al corazón (H4 y H3M1). También, el entrenamiento de resistencia produce un contenido total de LDH más bajo y una disminución en las formas de LDH de tipo muscular (H1M3, M4) (27).
Sin embargo, Newsholme (21) ha cuestionado el papel potencial del perfil de las isoenzimas de LDH en la producción vs. la utilización de La- en varios puntos: 1) la reacción de la LDH está cerca del equilibrio, lo que sugiere que la inhibición por piruvato tendría un efecto despreciable en el flujo a lo largo de la reacción, 2) las diferencias entre la función de las isoenzimas podrían ser menores in vivo que in vitro debido a la mayor temperatura y a otros factores del entorno intracelular 3) la inhibición por piruvato no se produciría en las elevadas concentraciones de la enzima LDH encontradas in vivo y 4) la inhibición informada, podría deberse a los rastros de la forma enólica del piruvato que son probablemente mas frecuentes in vitro que in vivo.
Además, Van Hall (27) recalcó lo siguiente: 1) que la cantidad absoluta de las formas de LDH del tipo corazón, permanece bastante constante entre los tipos de fibra y durante el entrenamiento de resistencia y 2) que las concentraciones de piruvato y La- necesarias para la inhibición de LDH in vitro son varias veces más altas que las concentraciones más altas observadas in vivo. También es posible que la unión de LDH a las proteínas celulares pueda cambiar sus parámetros cinéticos. Mi conclusión es que el papel regulador exacto de las isoenzimas de la LDH no ha sido completamente establecido todavía.
LDH: LOCALIZACION DENTRO DE LA CELULA
La lactato deshidrogenasa ha sido considerada durante mucho tiempo como una enzima glucolítica ' soluble ' localizada en el citoplasma de las células, predominantemente en la banda I del músculo estriado (18, 27). Además, existe evidencia significativa que LDH se une reversiblemente a las proteínas del citoesqueleto entre las que se incluyen, la actina, tubulina y troponina entre otras (ver (18) para obtener las referencias). Además, aparentemente un fragmento de LDH se localiza en el retículo sarcoplásmico (SR), particularmente en el músculo esquelético glucolítico (por ejemplo, (1)). De acuerdo con los estudios previamente citados, generalmente se informa que la actividad glucolítica se encuentra exclusivamente en el fragmento sobrenadante de los homogenatos de tejidos, fragmento que esencialmente no contiene ninguna mitocondria.
La historia de la localización de la LDH y por lo tanto, del metabolismo intracelular del La-, se volvió más cuestionada con una publicación de Baba y Sharma en 1971 (1). Los autores (1) citaron varios estudios que informaron actividad de la LDH en la fracción mitocondrial de los tejidos, así como numerosos estudios histoquímicos que demostraron la actividad de la LDH en la mitocondria de varios tejidos. Sus propios resultados (1), obtenidos en el músculo esquelético a través de una combinación de técnicas histoquímicas y de microscopía electrónica, indicaron principalmente la presencia de isoenzimas del tipo cardíaco en las mitocondrias y citoplasma, y principalmente isoenzimas del tipo-múscular en asociación con el retículo sarcoplásmico (SR). Los autores supusieron que las formas de LDH de tipo cardíaco, podrían desempeñar un papel similar al de la glicerol fosfato deshidridrogenasa y al de la malato deshidridrogenasa. Sin embargo, concluyeron que “la permeabilidad de la mitocondria al lactato no ha sido correctamente demostrada y el transporte de lactato sigue siendo pura especulación” ((1); el énfasis es mío).
Posteriormente, Skilleter y Kun (25), Deimann et al. (8), Kline et al. (16), Brandt et al. (3), y Chretien et al. (7) entre otros, a través de una variedad de técnicas, encontraron LDH en asociación con las mitocondrias. En 1972, Skilleter y Kun (25) también informaron la oxidación de La- por mitocondrias de hígado aisladas, aunque, increíblemente, el proceso parecía requerir energía. Szczesna-Kaczmarek (26) se transformó en el primer investigador que informó oxidación directa del La- por parte de las mitocondrias en el músculo esquelético en 1990.
EL TRANSPORTE INTRACELULAR DE LACTATO (LANZADERA DE LACTATO)
Finalmente, en 1999, Brooks et al. (5) esclarecieron totalmente el transporte de lactato intracelular que sólo había sido inferido por Baba y Sharma (1). Un principio central de este transporte intracelular fue que el La- es un producto inevitable de la glucolisis, especialmente durante la glucólisis rápida, ya que la LDH tiene la Vmax más alta que cualquier enzima de la vía glucolítica y la Keq de la reacción de conversión de piruvato a La- está desplazada hacia el La- y nicotinamida adenina dinucleotido(NAD+) (4,5). Dada esta información, Brooks et al (4) cuestionaron cómo sería posible que el lactato fuera transformado nuevamente en piruvato en el citosol, permitiendo así, la oxidación de La- por los tejidos bien perfundidos, un fenómeno universalmente aceptado, que sentó las bases de la teoría del transporte de lactato célula a célula (9, 10, 27). Una explicación simple sería que la LDH cataliza una reacción cercana al equilibrio, lo que permitiría que pequeños cambios en las concentraciones de sustratos y productos, dirijan inmediatamente la reacción en la dirección opuesta. Sin embargo, el laboratorio de Brooks (4, 5, 11, 13) en los últimos años, planteó una explicación diferente y presentó evidencia de los siguientes componentes claves de un transporte de lactato intracelular en el músculo esquelético: 1) captación y oxidación directa de La- por las mitocondrias aisladas sin conversión extra mitocondrial previa de La- a piruvato (Fig. 1), 2) la presencia de un conjunto de LDH intramitocondrial y 3) la presencia de un transportador de La-, transportador monocarboxilato 1 (MCT1), en la mitocondria, probablemente en la membrana interna mitocondrial.
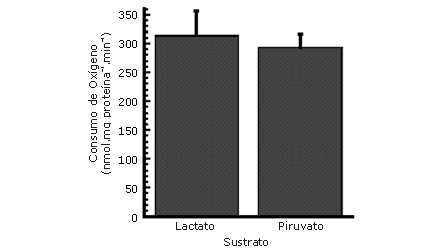
Figura 1. Oxidación de lactato y piruvato por mitocondrias aisladas de músculo esquelético de rata. La concentración de ambos sustratos fue de 10 mM y en el medio además se agregó malato en una concentración de 2,5 mM. Comparar con los valores informados por Yoshida et al. (30) en la Figura 2 quienes argumentan que los resultados que se muestran en esta figura se deben probablemente a contaminación. Los datos fueron extraídos de la Tabla 1 del trabajo de Brooks et al (5).
El funcionamiento de un transporte de lactato intracelular como el descripto por el grupo de Brooks (5) implica una producción constante de La- en el citosol, donde la tasa de producción aumenta cuando la actividad glucolítica es elevada. Debido a su elevada concentración en relación con el piruvato, el La- sería el principal monocarboxilato en difundir hacia la mitocondria. En la versión original de este transporte, este La- sería transportado luego a través de la membrana mitocondrial interna por MCT1. Una vez dentro de la matriz mitocondrial, la LDH mitocondrial catalizaría la reconversión de La- a piruvato quien se oxidaría, a través de la reacción de PDH a acetil coenzima A (CoA). Luego el acetil CoA ingresaría al ciclo de los ácidos tricarboxílicos. Un punto importante es que este transporte intracelular de lactato, no sólo aportaría el sustrato en forma de La- para la conversión al piruvato, si no que también aportaría los equivalentes reducidos (la forma reducida de NAD+ (NADH)), reemplazando o complementando así, el rol de los transportadores (lanzaderas) malato-aspartato y glicerol-fosfato en diferentes grados, dependiendo de la tasa de formación de La- y de su velocidad de transporte hacia la mitocondria (9).
Es correcta la teoría de Brooks? ¿Las mitocondrias pueden oxidar al La- directamente? Después de la propuesta de Brooks del transporte intracelular de lactato, Rasmussen et al. (23) y Sahlin et al. (24) no observaron ninguna evidencia que indicara que las mitocondrias puedan utilizar La- como un sustrato sin la conversión previa a piruvato en el citoplasma. Un informe preliminar de Willis et al. (29) también observó actividad insignificante del transporte intracelular de lactato propuesto en mitocondrias aisladas a partir de músculo esquelético (Tipos I y IIb) y de hígado de ratas. Ponsot et al. (22) no observaron ninguna señal directa de oxidación mitocondrial de La- en las fibras desolladas del músculo cardíaco, músculo esquelético glucolítico o músculo esquelético oxidativo. Finalmente, Yoshida et al. (30) del laboratorio de Bonen recientemente también observaron oxidación directa mínima de La- por las mitocondrias del subsarcolemma o por mitocondrias intermiofibrilares, ya sea del músculo esquelético rojo (oxidativo) o blanco (glucolítico) (Figura 2).
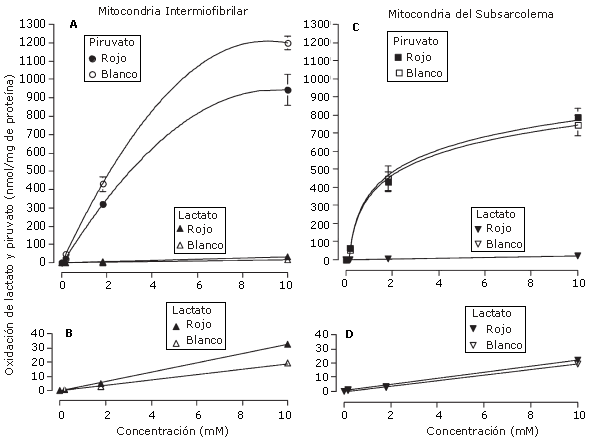
Figura 2. Oxidación de diferentes concentraciones (0,18, 1,8, y 10 mM) de lactato y piruvato en las mitocondrias intermiofibrilares (Panel A) y del subsarcolema (Panel C) de músculos rojos y blancos de ratas. La oxidación de lactato también se presenta en una escala 30 veces mas sensible en los dos tipos de músculos; rojo y blanco en las mitocondrias intermiofibrilares (Panel B) y del subsarcolema (Panel D). Es importante observar la oxidación insignificante directa de lactato en la mitocondrias intermiofibrilares y del subsarcolema obtenidas del músculo esquelético rojo y blanco de ratas. Comparar con los valores obtenidos por Brooks et al. (5) en la Figura 1. Hashimoto y Brooks (11) afirmaron que estos resultados se deben probablemente a la pérdida de LDH mitocondrial durante los procesos de aislamiento. (Obtenido de Yoshida, Y., G.P. Holloway, V. Ljubicic, H. Hatta, L.L. Spriet, D.A. Hood, y A. Bonen. Negligible direct lactate oxidation in subsarcolemmal and intermyofibrillar mitochondria obtained from red and white rat skeletal muscle. J. Physiol. 582:1317-1335, 2007. Copyright * 2007 Blackwell Publishing. Reproducido con autorización).
Un punto central en este debate es la localización exacta de la LDH. Aunque varios investigadores han informado que la LDH se asocia con las mitocondrias (como previamente describimos), estos informes deben ser discutidos con mayor detenimiento.
Primero, algunos investigadores (23, 24, 30) destacan que la cantidad de LDH asociada con las mitocondrias es demasiado pequeña y concluyen que las mitocondrias aisladas por Brooks et al. (5), así como por otros autores están contaminadas con LDH citoplásmica. De hecho, Chretien y colegas (7), quiénes encontraron LDH en preparaciones enriquecidas con mitocondrias de músculo esquelético humano, consideraron que la LDH era un contaminante del citoplasma. De manera contraria, Hashimoto y Brooks (11) apuntaron a diferencias en las técnicas de aislamiento de las mitocondrias y plantearon que otros autores probablemente perdieron la LDH que estaba asociada con las mitocondrias.
Claramente, sería útil que algún investigador comparara cuidadosamente la actividad de LDH y oxidación de La- en mitocondrias aisladas por los métodos de Brooks y colaboradores, en comparación con mitocondrias aisladas por las técnicas de otros investigadores que no hayan observado oxidación mitocondrial de La-.
Otro obstáculo crítico a la idea de oxidación de La- dentro de la matriz mitocondrial, es la posible violación de la primera ley de la termodinámica tal como lo describen Sahlin et al. (24). Las vías de entrada diferentes para el piruvato y NADH del citoplasma a la matriz mitocondrial, producen un marcado gradiente de potencial oxidación/reducción a través de la membrana interna mitocondrial. Para el mantenimiento de este gradiente es fundamental la existencia de un paso de no-equilibrio entre los transportadores (lanzaderas) de glicerol-fosfato y malato-aspartato. No se han presentado ni evidencias empíricas ni explicaciones teóricas que sustenten el paso de no-equilibrio en la hipótesis de transporte intracelular de lactato.
¿LDH EN EL ESPACIO INTERMEMBRANARIO MITOCONDRIAL?
En mi opinión, el peso de la evidencia está en contra de la presencia de LDH en la matriz mitocondrial y, por consiguiente, de la oxidación de La- dentro de la matriz mitocondrial. Sin embargo, la LDH podría estar presente en el espacio intermembranario de la mitocondria, quizás unida (laxamente?) a la membrana interna mitocondrial. Como mencionamos previamente, Baba y Sharma (1), encontraron LDH asociada con la mitocondria pero, al mismo tiempo, cuestionaron la permeabilidad de la mitocondria al lactato, dejando al espacio intermembranario como una ubicación probable en el músculo cardíaco y el músculo esquelético. Skilleter y Kun (25) realizaron fraccionamiento submitocondrial y llegaron a la conclusión que la LDH en la mitocondria intacta 'probablemente se encuentra en el lado exterior de la membrana interna ' en las células del hígado. Deimann et al. (8) utilizaron microscopía electrónica de transmisión y encontraron el producto de reacción para LDH ' claramente identificado en el espacio intermembranario de la mitocondria ' en músculo esquelético glucolítico de conejos. Utilizando ruptura por proteólisis de mitocondrias aisladas de hígado, Kline et al. (16) concluyeron que la LDH está 'principalmente en la membrana exterior y en el espacio periplásmico'. Brandt et al. (3) utilizaron fraccionamiento por digitonina de mitocondrias aisladas de corazón, riñón, hígado y linfocitos de rata; e informaron que 'la LDH mitocondrial se localiza principalmente en el espacio periplásmico'.
Espacio periplásmico y espacio intermembranario son términos equivalentes.
Observaciones recientes de Hashimoto et al. (12) del laboratorio de Brooks aportaron detalles adicionales sobre la posibilidad de que la LDH esté asociada con la membrana interna mitocondrial. Usando técnicas de microscopía electrónica de escaneo confocal láser e inmunoblotting luego de inmuno precipitación, en células L6 del músculo esquelético, Hashimoto et al. (12) encontaron evidencia que sugiere que LDH, MCT1, la glucoporteína transmembranaria que atraviesa una sola vez la membrana CD147 y la citocromo oxidasa, se colocalizan en la membrana mitocondrial interna, y la enzima LDH aparentemente se ubicaría en la superficie exterior de la membrana interna. Los autores (12) denominaron a éste complejo; complejo de oxidación del lactato y han descripto evidencia experimental que apoya la existencia de este complejo en otros trabajos (11). La ubicación potencial de la LDH unida laxamente al exterior de la membrana mitocondrial interna, replantea la posibilidad que dicha LDH podría ser susceptible a la pérdida durante el aislamiento de las mitocondrias del tejido muscular. Sin embargo, parecería improbable que la LDH (peso molecular igual a 134000) pueda atravesar una membrana mitocondrial exterior intacta porque la misma es impermeable a las moléculas superiores a 5000 daltons, mientras que las especies NAD+/NADH con un peso molecular de aproximadamente 664, la atraviesan fácilmente. También parecería improbable que la LDH dentro del espacio intermembranario mitocondrial sea susceptible a la destrucción por parte de las proteasas usadas en el aislamiento mitocondrial (tripsina con peso molecular de 23000; y nagarasa con un peso molecular de 27000).
EL TRANSPORTE INTRACELULAR DE LACTATO PERMANECE VIABLE
A pesar de las serias observaciones descriptas previamente, es bastante posible que opere un transporte intracelular de lactato, aunque sin un metabolismo de lactato dentro de la matriz (Figura 3).
Es razonable especular que las concentraciones de piruvato y NADH sean menores al lado de la membrana mitocondrial interna donde el transportador del piruvato y los transpordadores de NADH (malato-aspartato y glicerol fosfato) trasladan los equivalentes de piruvato y NADH, respectivamente, hacia adentro de la mitocondria. En otras palabras, las mitocondrias que realizan una oxidación activa producirían una 'disminución drástica ' en la utilización de piruvato y NADH y, por consiguiente, en su captación a partir de las ubicaciones citosólicas adyacentes. Al mismo tiempo, los sitios de glucólisis celular crearían concentraciones elevadas de La- dado que el principal producto final de la glucólisis es el La- debido a la elevada actividad de LDH, tal como lo describimos previamente. Esta situación produciría la mayor producción y concentración de La- en las localizaciones citosólicas alejadas de las mitocondrias. Luego, a causa de la concentración de La- relativamente superior en comparación con la concentración de piruvato, el La- sería la primera especie en difundir hacia las áreas cercanas a las mitocondrias. La concentración de La- es típicamente aproximadamente 10-200 veces mayor que la concentración de piruvato en el músculo esquelético. Al lado de las mitocondrias, o si Hashimoto et al. (12) están en lo cierto, en el espacio intermembranario, el La- y NAD+ serán transformados nuevamente a piruvato y NADH por la acción de la LDH, para ser incorporados a la mitocondria. Tal esquema, que se supone análogo al esquema del transportador de la fosfocreatina, concordaría con la cercanía al equilibrio de la reacción de la LDH en el citosol y daría lugar a la producción de La- junto con la oxidación subsecuente y menos transporte de La- hacia afuera de la célula.
Debo aclarar que la ubicación específica de la LDH, en el espacio intermembranario o unida a la membrana interna, no sería un requisito para el funcionamiento de un transportador de lactato intracelular. Finalmente, a pesar de la evidencia de la presencia de MCT1 en un complejo de oxidación de lactato (12), es bastante posible, quizás probable, que el piruvato sea transportado a través de la membrana interna mitocondrial a través del transportador mitocondrial de piruvato (MPC (14)) que tiene una afinidad muy alta por el piruvato.
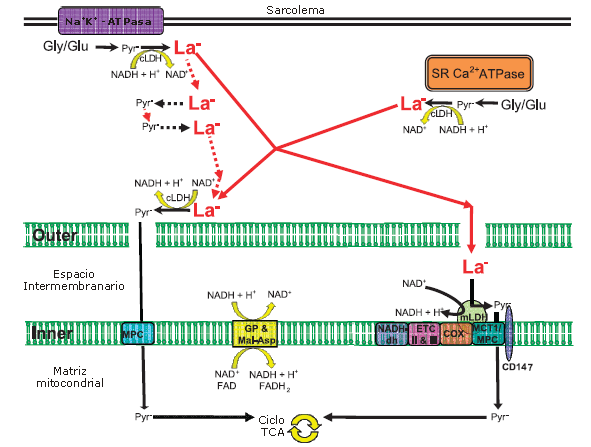
Figura 3. Representación esquemática de los elementos esenciales de los posibles transportadores (lanzaderas) de lactato intracelular (intramuscular), uno formando un complejo de oxidación de lactato con la lactato deshidridrogenasa (LDH) (12) en la membrana interna mitocondrial, y otro sin la LDH unida a la membrana interna o incluso en el espacio intermembranario. Se considera que una elevada actividad de LDH citosolica garantiza la formación de lactato (La-) en el citoplasma bajo virtualmente todas condiciones, pero sobre todo durante el ejercicio. Aunque el La- podría formarse en todo el citosol, se muestran dos situaciones particulares; una en asociación con la bomba Na+/K+-ATPasa en el sarcolema; y la otra, junto con la Ca2+-ATPasa en la membrana de SR. El sarcolema está representado por gruesas líneas dobles en la zona superior del esquema, mientras que las membranas mitocondriales internas y externas han sido agrandadas dramáticamente para demostrar las posibles vías del La-. Las líneas punteadas rojas y negras que conectan al Pyr- con el La- en el cuadrante izquierdo superior indican la naturaleza cercana al equilibrio de la reacción de la LDH y el movimiento de la masa de La- y Pyr- de un sitio de formación neta a un sitio de consumo neto. Los huecos en la membrana mitocondrial exterior ilustran que es libremente permeable a la mayoría de las moléculas pequeñas (pero probablemente impermeable a LDH, tripsina o nagarsa; ver el texto). El La- se presenta en negrita y rojo y de mayor tamaño que el piruvato (Pyr-) para indicar que el La- se presenta típicamente en mayor concentración que el Pyr-. Si se convirtiera nuevamente La- a Pyr- fuera del espacio intermembranario, dentro del espacio, o a través de una LDH mitocondrial, el NADH +H+ resultante sería transportado por la membrana interna mitocondrial a través de los transportadores (lanzaderas) malato-aspartato y glicrol fosfato. Es importante destacar que la ilustración de estos transportadores tradicionales en la membrana interna mitocondrial no pretende señalar que sus componentes se limitan a esta ubicación; las reacciones que se producen fuera de la membrana interna pueden producirse en todo el citosol. El Pyr- podría ser transportado a través de la membrana mitocondrial interna por un transportador de piruvato, (MPC, postulado como el más probable) o por un transportador monocarboxilato (MCT1), ya que ambos han sido identificados en la membrana interna. El MCT1/MPC en el complejo de oxidación de lactato no implica que Hashimoto et al. (12) informaran la presencia de MPC allí; su evidencia apoya el MCT1. Para evitar confusión, no se muestran las flechas que indican las posibles rutas de difusión de NAD+ y NADH + H+. COX = citocromo oxidasa; cLDH= lactato deshidridrogenasa citosólica; CD147= glicoporteína transmembranaria que atraviesa la membrana una sola vez; ETC II e III, complejos II y III de la cadena de transporte de electrones; Gly= Glucógeno; Glu=glucosa; Inner= membrana interna mitocondrial; La-= Lactato; MCT1=transportador monocarboxilato 1; mLDH= LDH mitocondrial; MPC= transportador de piruvato mitocondrial,; NADH-dh=NADH deshidridrogenasa complejo I; Outer= Membrana externa mitocondrial; Pyr-, Piruvato. [Adaptado de Hashimoto, T., R. Hussien, y G.A. Brooks. Colocalization de MCT1, CD147, y LDH en el mitocondrial la membrana interna de L6 músculo células: la evidencia de un mitocondrial lactato oxidación complejo. Es. J. Physiol. Endocrinol. Metab. 290:E1237y E1244, 2006. Copyright * 2006. The American Physiological Society Reproducido con autorización].
COMPARTIMENTALIZACION DEL METABOLISMO
El modelo previamente descrito es consistente con la compartimentalización del metabolismo como ha sido descripta en varios estudios. James y colegas (15) propusieron que la bomba Na+/K+ ATPasa obtenía su energía totalmente de la glucolisis quien está estrechamente asociada con la bomba; idea que ha sido corroborada recientemente con los estudios de fibras del músculo esquelético desolladas mecánicamente. Hay también evidencia considerable de que existe una compartimentalización funcional de la glucólisis con el SR. La Figura 4 proporciona la evidencia circunstancial visual de la compartimentalización glucolítica con el SR. ¿Es posible que el La- producido por glucólisis asociada con la bomba de Ca2+ del SR se desvíe hacia la mitocondria intermiofibrilar, mientras que el La- producido por la glucólisis asociada con la actividad de la bomba Na+/K+-ATPasa se desvíe hacia las mitocondrias del subsarcolema?. Quizás los estudios futuros proporcionarán la evidencia necesaria en éste aspecto.

Figura 4. Imagen de una fibra del músculo esquelético humano obtenida mediante microscopía electrónica de transmisión (TEM) (amplificación original 20000X; barra de la escala = 2 µm). La sección de 60 nm de espesor fue cortada precisamente en la superficie de la miofibrilla, para poder visualizar las tríadas situadas en la banda I a cada lado de la línea z, con largas mitocondrias apareadas localizadas transversalmente a nivel de la banda I alrededor del aparato contráctil y en el contacto con el SR pero claramente separadas de los túbulos t. Es posible observar rastros de túbulos longitudinales del SR como los cordones desde la cisterna terminal hacia el collar fenestrado al nivel de la banda M. Los abundantes gránulos de glucógeno se visualizan como puntos negros. Aunque algunos gránulos de glucógeno están indudablemente cerca de la mitocondria en la micrografía, la densidad de gránulos que rodean al SR es sugestiva de una compartimentalización glucolítica con el SR. Las fibras musculares humanas fueron fijadas con 2,5% glutaraldehido en buffer cacodilato de sodio 0,1 M (pH 7.3) durante 24 h luego fueron enjuagadas cuatro veces en buffer cacodilato de sodio 0,1 M, posteriormente fijadas en tetraóxido de osmio 1% (OsO4) y 1,5% ferrocianuro de potasio (K4Fe(CN)6) en buffer cacodilato de sodio 0,1 M durante 90 min, y enjuagadas dos veces en buffer cacodilato de sodio 0,1 M, deshidratadas con alcohol en diferentes graduaciones, infiltradas con mezclas graduadas de óxido de propileno y Epon, y embebidas en Epon al 100%. Las secciones ultra-delgadas (È60 nm) fueron cortadas con un ultramicrótomo Leica Supernova y contrastados con acetato de uranilo y citrato de plomo. Las secciones fueron examinadas y fotografiadas en un microscopio electrónico Philips EM 208 y una cámara Megaview III de FW. Las imágenes fueron registradas con una amplificación de 20000X con un microscopio electrónico de transmisión calibrado previamente. La figura fue obtenida por cortesía de Niels Krtenblad y Joachim Nielsen, de la Universidad de SouthernDenmark, Odense, Dinamarca. No ha sido publicada previamente.
CONCLUSIONES
En conclusión, las investigaciones sobre el metabolismo muscular del lactato siguen siendo un semillero de actividad 200 años después de que se informara que en los músculos que realizaban ejercicios, se producía un aumento de La-. La mayor fuente de discusión se centra en la vía/s intracelular exacta de oxidación de La-. El modelo de interés es el transporte intracelular del lactato. Las preguntas claves son las siguientes: ¿1) cómo se puede seguir el recorrido del La- de una localización intracelular a otra?; ¿2) dónde se localiza precisamente la LDH dentro de las células musculares?; ¿3) Para que funcione el modelo de transporte (lanzadera) intracelular del lactato es necesario que la LDH tenga localizaciones fijas? Los estudios futuros podrán proporcionar las respuestas conclusivas a estas preguntas.
Agradecimientos
El autor desea agradecer a Matthew L. Goodwin, Andrés Hernández, y James E. Harris por sus críticas y discusión de los proyectos del manuscrito.
Referencias
1. Baba N. and H. M. Sharma (1971). Histochemistry of lactic dehydrogenase in heart and pectoralis muscles of rat. J. Cell. Biol. 51:621-635
2. Benninga H (1990). A History of Lactic Acid Making: A Chapter in the History of Biotechnology. Boston, MA: Kluwer Academic Publishers, pp. 1-61
3. Brandt R. B., J. E. Laux, S. E. Spainhour and E. S. Kline (1987). Lactate dehydrogenase in rat mitochondria. Arch. Biochem. Biophys. 259: 412-422
4. Brooks G. A., M. A. Brown, C. E. Butz, J. P. Sicurello and H (1999). Dubouchaud. Cardiac and skeletal muscle mitochondria have a monocarboxylate transporter MCT1. J. Appl. Physiol. 87:1713-1718
5. Brooks G. A., H. Dubouchaud M. Brown, J. P. Sicurello and C. E. Butz (1999). Role of mitochondrial lactate dehydrogenase and lactate oxidation in the intracellular lactate shuttle. Proc. Natl. Acad. Sci. U.S.A. 96: 1129-1134
6. Cahn R. D., N. O. Kaplan, L. Levine and E. Zwilling (1962). Nature and development of lactic dehydrogenases. Science. 136:926-969
7. Deimann W., R. Freeman and H. D. Fahimi (1981). Improved contrast in cytochemistry of dehydrogenases by scanning transmission electron microscopy. J. Histochem. Cytochem. 29:678Y681
8. Gladden L. B (2004). Lactate metabolism: a new paradigm for the third millennium. J. Physiol. 558(Pt 1):5-30
9. Gladden L. B (2008). A lactatic perspective on metabolism. Med. Sci. Sports. Exerc. 40:477-485
10. Hashimoto T. and G. A. Brooks (2008). Mitochondrial lactate oxidation complex and an adaptive role for lactate production. Med. Sci. Sports Exerc. 40:486-494
11. Hashimoto T., R. Hussien and G. A. Brooks (2006). Colocalization of MCT1, CD147, and LDH in mitochondrial inner membrane of L6 muscle cells: evidence of a mitochondrial lactate oxidation complex. Am. J. Physiol. Endocrinol. Metab. 290:E1237-1244
12. Hashimoto T., S. Masuda, S. Taguchi and G. A. Brooks (2005). Immunohistochemical analysis of MCT1, MCT2 and MCT4 expression in rat plantaris muscle. J. Physiol. 567(Pt 1):121-129
13. James J. H., F. A. Luchette, F. D. McCarter and J. E. Fischer (1999). Lactate is an unreliable indicator of tissue hypoxia in injury or sepsis. Lancet. 354:505-508
14. Kline E. S., R. B. Brandt, J. E. Laux, S. E. Spainhour, E. S. Higgins, K. S. Rogers, S. B. Tinsley and M. G. Waters (1986). Localization of L-lactate dehydrogenase in mitochondria. Arch. Biochem. Biophys. 246:673-680
15. Nakae Y. and P. J. Stoward (1997). Effects of tissue protectants on the kinetics of lactate dehydrogenase in cells. J. Histochem. Cytochem. 45:1417-1425
16. Needham D. M., Machina Carnis (1971). The Biochemistry of Muscular Contraction in Its Historical Development. Cambridge, UK: Cambridge University Press, 782
17. Needham J (1970). Introduction. In: The Chemistry of Life; Eight Lectures on the History of Biochemistry, J. Needham (Ed.). Cambridge, UK: University Press, pp. vii-xxx
18. Newsholme E. A (2004). Enzymes, energy and endurance. Some provocative thoughts. In: Principles of Exercise Biochemistry, J.R. Poortmans (Ed.). Basel, Switzerland: Karger, pp. 1-35
19. Rasmussen H. N., G. van Hall and U. F. Rasmussen (2002). Lactate dehydrogenase is not a mitochondrial enzyme in human and mouse vastus lateralis muscle. J. Physiol. 541(Pt 2):575-580
20. Skilleter D. N. and E. Kun (1972). The oxidation of L-lactate by liver mitochondria. Arch. Biochem. Biophys. 152:92-104
21. Szczesna-Kaczmarek A (1990). L-lactate oxidation by skeletal muscle mitochondria. Int. J. Biochem. 22:617-620
22. Van Hall G (2000). Lactate as a fuel for mitochondrial respiration. Acta. Physiol. Scand. 168:643-656
23. A Biographical Dictionary of Scientists (1982). 3rd ed. T.I. Williams (Ed.). New York, NY: John Wiley and Sons
24. Willis W. T., A. Thompson, J. I. Messer and J. S. Thresher (2003). Vmax of mitochondrial electron shuttles in rat skeletal muscle and liver (Abstract). Med. Sci. Sports. Exerc. 35:S396
25. Yoshida Y., G. P. Holloway, V. Ljubicic, H. Hatta, L. L. Spriet, D. A. Hood and A. Bonen (2007). Negligible direct lactate oxidation in subsarcolemmal and intermyofibrillar mitochondria obtained from red and white rat skeletal muscle. J. Physiol. 582(Pt 3):1317-1335
Cita Original
Gladden, L.B. 200th anniversary of lactato research in muscle. Exerc. Sport Sci. Rev., Vol. 36, No. 3, pp. 109-115, 2008.
Cita en PubliCE
Bruce L Gladden (2012). Bicentenario de las Investigaciones sobre el Lactato en el Músculo. .https://g-se.com/bicentenario-de-las-investigaciones-sobre-el-lactato-en-el-musculo-1277-sa-a57cfb271e1def