Altura y Entrenamiento de Resistencia
Heikki Rusko1,4, Heikki Tikkanen1,2,3, Juha Peltonen1,21 KIHU - Research Institute for Olympic Sports Jyväskylä Finland
2 Unit for Sports and Exercise Medicine , Institute of Clinical Medicine University of Helsinki , Helsinki, Finland
3 Folkhälsan Research Centre Biomedicum Helsinki Helsinki Finland
4 Department of Biology of Physical Activity University of Jyväskylä Jyväskylä , Finland
Resumen
Los beneficios de vivir y entrenar en altitud (HiHi) para lograr un mejor rendimiento deportivo en altitud están claros, pero los estudios controlados sobre la mejora del rendimiento al nivel del mar son controversiales. Las razones por las cuales el entrenamiento en altitud (HiHi) no tiene efectos positivos incluyen: (1) el efecto de aclimatación podría haber sido insuficiente para que los atletas de élite estimulen un aumento en la masa de glóbulos rojos/masa de hemoglobina debido a una altura demasiado baja (menos de 2000-2200 m) y/o también a que el período de entrenamiento en altura fue demasiado corto (inferior a 3-4 semanas); (2) los efectos del entrenamiento en altura pueden haber sido afectados por estímulos de entrenamiento insuficientes para mejorar la función neuromuscular y del sistema cardiovascular; y (3) mayor estrés con posibles síntomas de sobre entrenamiento y una mayor frecuencia de infecciones. Por otra parte, los efectos de la hipoxia en el cerebro pueden influir en la intensidad del entrenamiento y en las respuestas fisiológicas durante el entrenamiento en altitud. Así, interrumpir la exposición a hipoxia entrenando en normoxia puede ser un factor importante para evitar o minimizar los efectos nocivos asociados a la hipoxia crónica. Al comparar HiHi y HiLo (vivir en altitud y entrenar en baja altitud), es obvio que las dos condiciones pueden inducir un efecto de aclimatación positivo y pueden aumentar la capacidad de transporte de oxígeno de la sangre, por lo menos en los sujetos “respondedores”, si se cumplen ciertos requisitos previos. La dosis mínima para lograr un efecto de aclimatación hematológica debe ser mayor a 12 h por día durante por lo menos 3 semanas en altura o en altitud simulada de 2100-2500 m. La exposición a hipoxia tendría algunos efectos de transferencia positivos en el entrenamiento subsiguiente en normoxia durante y después de HiLo. La mayor capacidad de transporte sanguíneo de oxígeno permite realizar entrenamiento de mayor intensidad durante y después de HiLo en condiciones de normoxia subsiguientes, lo que incrementa el potencial para mejorar algunos determinantes neuromusculares y cardiovasculares de rendimiento de resistencia. Los efectos del entrenamiento en hipoxia y de la hipoxia severa intermitente a corto plazo en reposo no están todavía claros y deben ser investigados con detalle.
Palabras Clave: atletas, función cardiovascular, eritropoyetina, entrenamiento en hipoxia, hipoxia intermitente, consumo de oxígeno máximo, masa de eritrocitos
INTRODUCCION
En los últimos 20-30 años, se ha incrementado el consumo de oxígeno máximo (VO2max) de los atletas y los records mundiales en carreras de fondo han mejorado. Uno de los factores que influyen en el aumento en el VO2max y en la mejora de los records mundiales es el entrenamiento en altitud, porque los atletas que viven en las regiones montañosas han ganado muchas medallas en las Olimpíadas y en los Campeonato Mundiales de carreras de distancia en las últimas décadas. Sin embargo, la literatura científica sobre el entrenamiento en altitud de sujetos que habitan al nivel del mar es confusa, y la mayoría de los estudios controlados no han observado un efecto positivo en el rendimiento a nivel del mar. Esto ha fomentado la búsqueda de estrategias alternativas para usar la hipoxia como un estímulo adicional para atletas. Durante los últimos 10 años, han aumentado notablemente la cantidad de técnicas y métodos diseñados para simular la hipoxia generada por la altitud y el entrenamiento en altitud. Se han desarrollado alojamiento en lugares de elevada altitud y cámaras de altitud, así como también equipos de respiración especiales para aportar una inspiración hipóxica en reposo y durante el ejercicio, y se ha creado el término “entrenamiento en hipoxia intermitente” (IHT). El entrenamiento en hipoxia intermitente hace referencia al uso discontinuo de hipoxia normobárica o hiperbárica, en un esfuerzo por aprovechar los beneficios del entrenamiento en altitud natural y evitar los problemas relacionados al mismo. En general, los entrenamientos en hipoxia intermitente pueden ser divididos en dos estrategias diferentes: (1) utilizando hipoxia en reposo, con el objetivo principal de favorecer la aclimatación a la altitud o (2) generando hipoxia durante el ejercicio, con la meta principal de reforzar el estímulo de entrenamiento (Levine, 2002).
En esta revisión, abordaremos los problemas relacionados a la altitud, a la hipoxia y entrenamiento de resistencia en relación a los deportes competitivos. Esta revisión intenta establecer los efectos (positivos o negativos) del entrenamiento en altitud (vivir alto y entrenar alto o HiHi) en atletas que viven a nivel del mar. Además, discutiremos los beneficios y problemas de las diferentes metodologías de IHT, es decir, vivir alto y entrenar bajo (HiLo), exposición a hipoxia severa intermitente a corto plazo en reposo (IHR a corto plazo), y vivir bajo y entrenar en hipoxia (entrenamiento en hipoxia, HT), con especial referencia al rendimiento a nivel del mar. Si no podemos encontrar datos de atletas de elite o bien entrenados, utilizaremos los datos provenientes de individuos activos físicamente o de individuos desentrenados.
RENDIMIENTO DE RESISTENCIA Y RESPUESTA DE ENTRENAMIENTO AL NIVEL DEL MAR Y EN ALTITUD
Los estímulos que inducen los efectos de entrenamiento en atletas de resistencia pueden ser divididos en mecánicos, neuromusculares y químicos (por ejemplo fuerza, tensión, actividad neural, hormonas, tensión de oxígeno, para mas ejemplos consultar Rusko 2003). En condiciones de altitud, se sabe que la exposición a hipoxia influye en todos los sistemas funcionales del cuerpo, entre los que se incluyen el sistema nervioso central, sistema respiratorio, sistema cardiovascular y muscular. A nivel tisular, la hipoxia promueve un rápido censado del oxígeno y las posteriores funciones celulares. El modelo actual se basa en el factor inducible por hipoxia 1 (HIF-1) que está compuesto por una subunidad α y una subunidad β. En normoxia, HIF-α se degrada rápidamente. En hipoxia, los HIF-α recientemente formados son estabilizados y translocados hacia el núcleo y son dimerizados con subunidades de HIF-β, que están presentes independientemente de la presión parcial de oxígeno (PO2) para formar HIF-1. El HIF-1 se une a los elementos sensibles a hipoxia de los genes blanco de HIF-1 y estimula la síntesis de la proteína correspondiente (por ejemplo eritropoyetina, eritropoyesis; el factor de crecimiento endotelial vascular, angiogénesis; óxido nítrico sintetasa, vasodilatación; enzimas glucolíticas, metabolismo anaeróbico; hidroxilasa de tirosina, síntesis de catecolaminas) Qelkmann y Hellewig-Bürgel, 2001,Lahiri et al, 2002). Por lo tanto, vivir y entrenar en hipoxia puede influenciar casi todos los órganos y tejidos del cuerpo. Sin embargo, los efectos principales de las diferentes metodologías de entrenamiento en altitud pueden observarse en aquellos órganos y tejidos que influyen en el rendimiento aeróbico y anaeróbico.
En los atletas de resistencia, la capacidad de los músculos para recibir y consumir oxígeno excede la capacidad del sistema cardiovascular para transportarlo (Wagner, 2000). El objetivo principal del entrenamiento en altitud es aumentar el volumen total de glóbulos rojos y de hemoglobina para mejorar el vinculo limitante (es decir, el suministro de oxígeno) aumentando la capacidad de la sangre arterial para transportar oxígeno, y de este modo aumentar el VO2max y mejorar el rendimiento, tanto a nivel del mar como en altitud. El entrenamiento en altitud también puede inducir algunos efectos negativos sobre los determinantes de rendimiento (Rusko, 1996; Bailey y Davies, 1997; Hahn y Gore, 2001), enmascarando y anulando los beneficios del aumento de glóbulos rojos. Una posible razón de la ausencia de efecto positivo del entrenamiento en altitud es que, incluso una hipoxia moderada durante el ejercicio puede comprometer sustancialmente el ritmo del entrenamiento y disminuir los estímulos mecánicos y neuromusculares, lo que provoca un debilitamiento gradual de algunos determinantes específicos del rendimiento de resistencia. Por lo tanto, aunque el VO2max y el transporte de oxígeno desempeñan un papel fundamental en la mayoría de los deportes de resistencia, es necesario tener en cuenta otros factores, como las características neuromusculares y anaeróbicas ("factores de potencia muscular”), que influyen en el rendimiento de resistencia y en la respuesta al entrenamiento frente al entrenamiento en altitud .
Varios estudios han demostrado que, en condiciones de normoxia, las características neuromusculares y anaeróbicas pueden diferenciar grupos homogéneos de corredores en función de su rendimiento en carreras de distancia (Bulbulian et al, 1986; Rusko y Nummela, 1996; Paavolainen et al, 1999a, b). Algunos estudios de “puesta a punto” han observado una mejora en el rendimiento de resistencia con entrenamiento reducido sin un cambio en el VO2max (Shepley et al, 1992; Houmard et al, 1994). Los corredores también alcanzaron un mayor VO2max durante la carrera en ascenso que durante la carrera en el llano y mientras más cerca estaba el VO2max de la carrera horizontal del VO2 de la carrera en ascenso, mayor era la relación con un mejor rendimiento en carrera (Paavolainen et al, 2000). Por otra parte, la activación previa de los músculos antes de realizar contacto con el suelo durante la carrera, que se sabe que está pre-programada en los centros superiores del sistema nervioso central, es mayor y el tiempo de contacto es menor en los corredores de alto calibre que en los corredores de menor calibre, con similares VO2max (Paavolainen et al, 1999b). También se ha demostrado que el entrenamiento de la fuerza mejora el rendimiento de los atletas de resistencia sin producir cambios en el VO2max (Johnston et al, 1997; Paavolainen et al, 1999c).
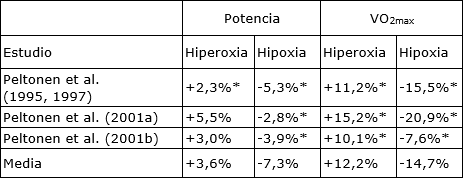
Por otra parte se han estudiado los factores limitantes de la capacidad de resistencia y el rol del oxígeno suministrado durante el ejercicio a diferentes alturas o en diferentes fracciones de oxígeno del aire inspirado (FiO2). Los efectos de la hipoxia aguda y crónica y la hiperoxia en el rendimiento físico han sido recientemente revisados por Noakes et al. (2001). Tanto en condiciones de hipoxia como de hiperoxia, el cambio en VO2max supera al cambio en el rendimiento físico, lo que sugiere que algunos factores influyen en el rendimiento físico independientemente de los cambios en el VO2max (Tabla 1).
A pesar de que el rendimiento físico y el VO2max no varían de manera similar con los cambios en FiO2, existe una fuerte correlación positiva entre los cambios en la carga de trabajo máxima y el gasto cardíaco máximo tanto en hipoxia como en hiperoxia (Peltonen y otros, 2001). Curiosamente, el gasto cardíaco máximo de atletas de élite sería menor durante la hipoxia aguda en comparación con la normoxia, a pesar de que existe una gran necesidad de aumentar el gasto cardíaco, tal como se observa por el mayor gasto cardíaco durante el ejercicio submáximo en hipoxia (Peltonen y col., 2001) y la activación simpática máxima (Lundby et al, 2001a).
Tabla 1. La producción de potencia máxima y el VO2max en hipoxia e hiperoxia agudas en comparación con normoxia indica cambios más pequeños en la potencia que en el VO2max. * Presenta diferencias significativas con la condición de normoxia, P <0,05.
También se ha demostrado que el rendimiento de resistencia en normoxia e hipoxia depende de la función del sistema neuromuscular y no sólo del suministro y del agotamiento de la energía (Kayser et al, 1994; Peltonen et al, 1995, 1997; Paavolainen et al, 1999b;. Kay et al, 2001; Carter et al, 2003, para más referencias, ver Noakes, 2000; Noakes et al, 2001). Durante el ejercicio máximo en hipoxia, tanto la actividad eléctrica muscular como el gasto cardíaco máximo se atenúan (Kayser et al, 1994; Peltonen et al, 1997) en comparación con lo que ocurre en condiciones de normoxia e hiperoxia, lo que sugiere que el sistema nervioso central desempeña un papel en la limitación del rendimiento físico. Por lo tanto, el menor gasto cardiaco máximo y el menor VO2max en hipoxia pueden ser el resultado y no la causa de la reducción en el reclutamiento del músculo esquelético, lo que apoya la existencia de una relación controlada (o "gobernador central") entre la función cardiaca y el rendimiento muscular (Peltonen y col, 2001; véase también Noakes et al, 2001). No está claro si el supuesto "gobernador central" es el responsable de estas respuestas o si simplemente indican que la depresión neuronal y cerebral en hipoxia no es un proceso regulado. Existe cada vez mas evidencia de una menor respuesta del sistema nervioso central inducida por contracciones musculares fatigantes (Gandevia, 2001; Di Lazzaro et al, 2003).
En conclusión, el rendimiento físico, el VO2max y la respuesta al entrenamiento a nivel del mar y en altitud no dependerían solamente de la entrega y utilización del oxígeno, sino que también de otros factores relacionados con la capacidad del sistema nervioso central para reclutar los músculos. De acuerdo con la hipótesis del gobernador central, el sistema nervioso central decide cuánto músculo debe reclutarse y cuanta fuerza y potencia deben producirse; y por lo tanto regula por ejemplo la función de bombeo del corazón. Probablementee, tanto los mecanismos de retroalimentación (por ejemplo, quimiorreceptores, barorreceptores, propioceptores) como los de estimulación podrían afectar la función del sistema nervioso central. Con el entrenamiento óptimo, el mayor reclutamiento continuo de los músculos provoca nuevos valores de gasto cardíaco máximo, VO2max y rendimiento físico de mayor nivel (ver Rusko 2003).
EJERCICIO EN HIPOXIA/EN ALTITUD
La saturación de oxígeno de la hemoglobina arterial (SaO2%) disminuye en la altura y además se sabe que algunos atletas de resistencia de elite desarrollan hipoxemia arterial inducida por el ejercicio durante el ejercicio máximo, incluso a nivel del mar (Dempsey et al, 1984; Dempsey y Wagner, 1999). La pequeña disminución adicional en SaO2%, incluso a bajas altitudes disminuye el VO2max en deportistas de élite a aprox. 600 m (VO2max aprox. 5,51 min-1 y aprox. 77 ml kg-1 min-1; Gore et al, 1996), en atletas bien entrenados a aprox. 900 m (VO2max aprox. 5,01 min-1, aprox. 72 ml kg-1 min-1; Terrados et al, 1985), en los atletas recreacionales a 1200 m (VO2max aprox. 4,41 min-1, aprox. 60 ml kg-1 min-1; Squires y Buskirk, 1982) y en atletas mujeres de élite a aprox. 600 m (aprox. 3,61 min-1, aprox. 61 ml kg-1 min-1; Gore et al, 1997). La disminución del VO2max es mayor cuanto mayor sea la altitud y cada disminución de 1% en SaO2% por debajo del nivel de 95% se aproxima a una reducción de 1-2% en VO2max (Dempsey y Wagner, 1999). En consecuencia, en hipoxia los atletas están obligados a entrenar en un VO2max inferior y con menores velocidades de entrenamiento máximo sostenido y de entrenamiento intervalado que en condiciones de normoxia (Gore et al, 1996, 1997; Brosnan et al, 2000). En hipoxia, la SaO2% de los deportistas de élite también disminuye durante el ejercicio submáximo (60-85% VO2max) (Gore et al, 1995; Peltonen et al, 1999, 2001). En consecuencia, en condiciones de hipoxia los atletas están obligados a entrenar a velocidades de entrenamiento submáximas más bajas (Levine y Stray-Gundersen, 1997; Hahn et al, 2001) lo que puede disminuir aún más los estímulos físicos y neuronales para los músculos en comparación con el entrenamiento en condiciones de normoxia. Como indicación de un menor estímulo de entrenamiento neuromuscular, se observó una reducción en la actividad en el electromiograma integrado (iEMG) en comparación a lo que ocurre en normoxia durante el ejercicio máximo en hipoxia crónica (Kayser et al., 1994) y aguda (Peltonen y col., 1997). Sin embargo, la actividad iEMG aumenta con la administración de oxígeno en altitud (Kayser et al., 1994).
La hipoxia sostenida provoca una reducción ampliamente conocida en el gasto cardíaco máximo (para mas referencias, consultar Hahn y Gore, 2001) y, contrariamente a las conclusiones que se plantearon anteriormente, el gasto cardíaco máximo puede disminuir durante la hipoxia aguda, al menos en atletas de resistencia de elite (Peltonen et al., 2001). También hay cada vez mas evidencia de que tanto el gasto cardíaco máximo como la frecuencia cardíaca máxima (FCmáx) disminuyen durante el ejercicio máximo en hipoxia (Peltonen et al., 2001; para más referencias, consultar Hahn y Gore, 2001). La menor altitud para la disminución de la FCmáx estaría cerca de 3100 m, y por encima de ella la FCmáx será menor cuanto mayor sea la altitud (Lundby et al., 2001b). La disminución de la FCmáx se relaciona de manera lineal con la disminución en SaO2% (Benoit et al., 2003) y, por lo tanto, los atletas de élite con mayor disminución de SaO2% en altitud pueden experimentar disminución en la FCmáx incluso en altitudes <3100 m. Similar a lo que ocurre con la actividad iEMG, la FCmáx aumenta con la administración de oxígeno acercándose a los valores del nivel del mar tanto en hipoxia aguda como en hipoxia crónica (Lundby et al., 2001b) (Kayser et al, 1994;. Boushel et al., 2001), y el gasto cardíaco máximo es mayor en hiperoxia aguda que en hipoxia (Peltonen et al, 2001) o luego de la administración de oxígeno en hipoxia crónica (Boushel et al, 2001).
La hipoxia durante el ejercicio desplaza hacia la izquierda las curvas de ventilación, frecuencia cardíaca, ritmo cardíaco y concentración de lactato sanguínea versus velocidad en comparación con los valores obtenidos a nivel del mar (Figura 1). Además, la curva de concentración de lactato sanguíneo en función de la frecuencia cardíaca se desplaza hacia la izquierda, especialmente a frecuencias cardiacas elevadas y, como consecuencia, la frecuencia cardíaca de entrenamiento puede disminuir ligeramente durante el entrenamiento en hipoxia (Levine y Stray-Gundersen, 1997; Peltonen et al, 1999, 2001; Hahn et al, 2001). Del mismo modo, el VO2 sub-máximo en una frecuencia cardiaca dada es mucho menor en hipoxia (Peltonen et al, 2001; Figura 1), y debido a que durante el ejercicio submáximo el volumen de plasma disminuye y la frecuencia cardiaca se incrementa, el volumen sistólico y el gasto cardíaco pueden también disminuir durante el entrenamiento de resistencia en altitud (Wolfel et al, 1991; Peltonen et al, 2001) a pesar de que la curva de gasto cardíaco en función del VO2 se haya desplazado hacia la izquierda (Peltonen et al, 2001;. Figura 1).
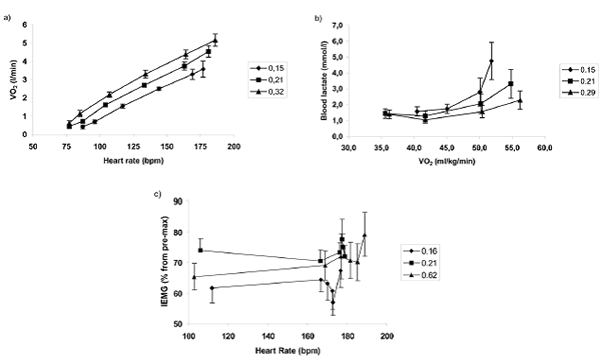
Figura 1. Cambios en (a) curvas de VO2 en función de la frecuencia cardiaca durante el ejercicio incremental hasta el agotamiento (modificado de Peltonen y col., 2001), (b) curvas de concentración de lactato sanguínea versus VO2 durante ejercicio submáximo incremental (modificado de Peltonen y col., 1999), y (c) curvas de actividad iEMG en función de la frecuencia cardíaca durante una prueba contrarreloj de remo máxima de 2500 m (modificado de Peltonen y col., 1997. Los puntos de datos indican valores registrados en el inicio y en intervalos de 500 m durante la prueba contrarreloj) en condiciones de hipoxia, normoxia e hiperoxia agudas. En hipoxia, el VO2 en la misma frecuencia cardíaca submáxima y concentración de lactato sanguínea es menor, y el reclutamiento de los músculos durante el ejercicio sostenido es menor, que en condiciones de normoxia o hiperoxia. Para una explicación más detallada consultar el texto. Los símbolos indican la fracción de oxígeno en el aire inspirado: ♦, 0,15; ■, 0,21; ▲, 0,32.
En resumen, hay muchas razones posibles para la ausencia de un efecto de entrenamiento positivo en altitud y una de estas razones es que incluso la hipoxia moderada durante el entrenamiento, puede comprometer sustancialmente el ritmo de entrenamiento y disminuir los estímulos mecánicos y neuromusculares que permiten alcanzar un efecto de entrenamiento. Además, hallazgos recientes sugieren que los estímulos de entrenamiento para el sistema cardiovascular y para el consumo de oxígeno también pueden disminuir durante el entrenamiento en hipoxia, lo que provoca un debilitamiento gradual de algunos determinantes específicos del rendimiento de resistencia.
COMPARACIÓN DE HIHI Y HILO EN LA INDUCCIÓN DEL EFECTO DE ACLIMATACIÓN
Los efectos de la aclimatación se han estudiado en entornos naturales durante la exposición continua y durante el entrenamiento en altitud, cuando se viaja ida y vuelta entre sitios de mayor y menor altitud, y en entornos artificiales utilizando hipoxia hipobárica o normobárica o en el nivel del mar. Son pocos los sitios naturales para llevar a cabo HiLo, pero permiten una exposición a hipoxia prolongada diaria (>20 h diarias). Una posibilidad de aplicación de HiLo es para aumentar la concentración de oxígeno del aire inspirado durante el entrenamiento en altitud natural (Chick et al., 1993). Otra metodología que se utiliza es la llamada "casa de altitud”, en la cual se crea una atmósfera de vida hipóxica normobárica a nivel del mar a través de la disminución al 15-16% de oxígeno en la concentración de oxígeno de una casa, apartamento o tienda de campaña, lo que equivale a una altitud de aprox. 2500 m y una presión parcial de oxígeno inspirado de 110 a 120 mmHg (Rusko et al, 1995a; Rusko, 1996). Una casa de altitud permite que las personas permanezcan en hipoxia normobárica de 12-20 h sin limitar la vida normal o los esquemas de entrenamiento a nivel del mar. Por el contrario, cuando se utilizan cámaras hipobáricas, el tiempo de exposición diaria a hipoxia es por lo general de sólo unas pocas horas y la diferencia de presión limita las transiciones entre la hipoxia y normoxia. Una revisión reciente sintetiza las diferentes técnicas para modificar la presión parcial del oxígeno inspirado (Wilber, 2001).
Independientemente de la metodología, una presión parcial reducida del oxígeno inspirado provoca una reducción inmediata de la presión parcial de oxígeno en la sangre arterial (PaO2), así como también en SaO2% si la caída de la PaO2 es lo suficientemente grande. Después de la inmediata caída inicial durante los primeros minutos de hipoxia, se ha observado un ligero aumento en la SaO2% utilizando oximetría de pulso (SpO2%) durante una noche en hipoxia; sin embargo, durante noches consecutivas en condición de hipoxia, la SpO2% nocturna parece estabilizarse y se mantiene en un nivel más bajo (Hahn et al., 2001).
La disminución de la SaO2% induce un aumento en la liberación renal de eritropoyetina a las pocas horas. Independientemente del método por el cual se introduce la hipoxia, el aumento en la concentración sanguínea de eritropoyetina es mayor cuanto mayor es la hipoxia y mayor es la disminución de la SaO2% (por ejemplo Eckhardt et al, 1989;. Berglund, 1992; Piehl-Aulin et al, 1998). La altitud umbral para estimular una liberación de eritropoyetina sostenida va desde 2100 m hasta 2500 m (Ri-Li et al., 2002). Notablemente, la concentración matutina de eritropoyetina se mantiene en un nivel más alto después de 2-5 días durante HiLo que durante HiHi (Figura 2). Sin embargo, en condiciones normales de oxigeno durante las horas del día en HiLo, la concentración de eritropoyetina disminuye significativamente en comparación con los valores obtenidos temprano por la mañana (Rusko et al, 1995a; Stray-Gundersen et al., 2000), debido a la corta hemivida de eliminación de la eritropoyetina (Berglund, 1992; Jensen et al, 1995).
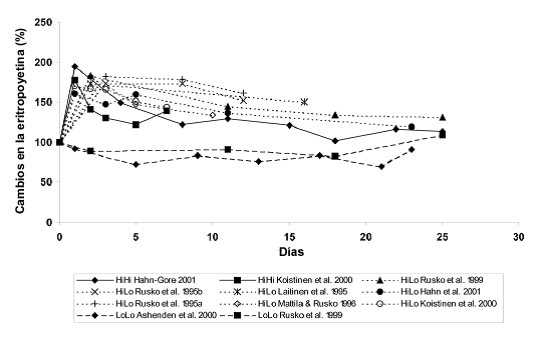
Figura 2. Cambios porcentuales en la concentración sérica de eritropoyetina (EPO) durante HiHi, HiLo y LoLo. Los valores medios de los diferentes estudios sugieren que durante HiLo, el nivel de eritropoyetina es mayor que durante HiHi después de 2-4 días.
Para entender la naturaleza transitoria del aumento en la concentración de eritropoyetina y las diferencias entre la exposición a HiHi y HiLo, es necesario tener en cuenta que la concentración sérica de eritropoyetina refleja un equilibrio entre la producción de eritropoyetina en los riñones y el consumo de la misma por la médula ósea. Al arribar a altitud, la producción de eritropoyetina es más alta que el consumo por lo que la concentración de eritropoyetina sérica aumenta. Sin embargo, el aumento en la actividad eritroide provoca un mayor consumo de eritropoyetina, lo que produce una caída en la concentración de eritropoyetina. Luego se establece un nuevo equilibrio dinámico con un leve aumento en la concentración sérica de eritropoyetina que se manifiesta por una elevada tasa de recambio de eritropoyetina. Las mediciones de la concentración sérica de eritropoyetina por sí solas no reflejarán este aumento sostenido en la producción de eritropoyetina que mantiene un mayor nivel de eritropoyesis (Grover y Bartsch, 2001, pp. 501-502).
Para obtener un efecto de aclimatación por la exposición a la hipoxia, el aumento en la producción de eritropoyetina debe producir un aumento en la capacidad de transporte de oxígeno de la sangre que se puede medir por el volumen total de glóbulos rojos y/o por la masa de hemoglobina. Los estudios previos de los participantes no entrenados indican que el volumen total de glóbulos rojos se incrementa en los habitantes de altitud en comparación con las personas que viven a nivel del mar, y la presión parcial crítica de oxígeno en la sangre arterial que produce un mayor volumen total de glóbulos rojos es de alrededor de 65 mmHg, lo que corresponde a una altitud entre 2000 y 2600 m (Reynafarje et al, 1959; Weil et al, 1968;.Fiori et al, 2000; Böning et al, 2001; Schmidt et al., 2002). A mayor altitud, el volumen total de glóbulos rojos es mayor cuanto mayor sea la altitud de residencia. En los sujetos que viven a nivel del mar, la aclimatación durante 3-4 semanas aumenta significativamente el volumen total de glóbulos rojos de individuos no entrenados incluso sin realizar entrenamiento en altitud (e.g. Reynafarje et al., 1959; Wolfel et al., 1991; Heinicke et al., 2003). Es difícil aplicar estos estudios a los atletas de resistencia que tienen un volumen total de glóbulos/masa de hemoglobina y volumen sanguíneo 20-25% superiores que los individuos no entrenados, incluso sin realizar entrenamiento en altitud (Remes, 1979; Böning et al., 2001; Schmidt et al, 2002); Sin embargo, los estudios transversales indican que existe un efecto acumulativo de la exposición a la altitud y del entrenamiento en altitud (Schmidt et al., 2002).
Muchos estudios longitudinales también indican que vivir alto y entrenar alto (HiHi) en alturas ≥2500 m durante 4 semanas aumenta significativamente el volumen total de glóbulos rojos/masa de hemoglobina en los atletas (Stray-Gundersen et al, 1992, 1993, 1995; Levine y Stray-Gundersen, 1997). Se ha observado un aumento del 6% en el volumen de glóbulos rojos en 14 corredores de fondo de élite después de 4 semanas de entrenamiento en altura de aprox. 2200m utilizando un método confiable (método de dilución de isótopos, con glóbulos rojos marcados con tecnecio) para medir el volumen total de glóbulos rojos (Rusko y Tikkanen, 1996). Se ha calculado que el aumento de la masa de hemoglobina durante el entrenamiento en altura es bastante pequeño y lento (aprox. 1% por semana; Berglund 1992) y que el volumen total de glóbulos rojos puede seguir aumentando durante 6-8 meses con la exposición a la altura (Reynafarje et al., 1959). Algunos estudios no han logrado observar aumentos en el volumen total de glóbulos rojos/masa de hemoglobina en los atletas después de HiHi. Gore et al. (1998) observaron que la masa de hemoglobina se mantenía sin cambios después del entrenamiento a 2690 m durante 31 días, a pesar de que todos los participantes tenían síntomas clínicos (véase la sección siguiente). En otro estudio realizado con esquiadores de fondo de élite, el entrenamiento durante 1 mes a aprox. 1900 m provocó un aumento en el volumen de sangre (+7%) y en la masa de hemoglobina (+3%), pero los cambios no fueron significativos (Svedenhag et al., 1997).
Algunos estudios revelaron que con la metodología HiLo (vivir en una altura de aprox. 2500 m y entrenar en alturas mas bajas de aprox. 1300 m) durante 4 semanas, se produjo un aumento aprox. un 5% en el volumen total de glóbulos rojos/masa de hemoglobina (Levine et al, 1991; Levine y Stray-Gundersen, 1997; Wehrlin et al, 2003). De manera similar, los estudios con HiLo que utilizaron una casa situada a nivel del mar (Laitinen et al, 1995; Rusko et al, 1999) observaron un aumento significativo de aprox. 5% en el volumen total de glóbulos rojos (glóbulos rojos marcados con tecnecio) o de masa de hemoglobina (método de re-inhalación de CO) en atletas de resistencia de elite durante el entrenamiento de 3 a 4 semanas. En un estudio, realizado en una casa de altura, el aumento de 3,5% en la masa de hemoglobina después de 4 semanas de HiLo no fue estadísticamente significativo (Piehl-Aulin, 1999). En algunos estudios HiLo, se observaron concentraciones matutinas de eritropoyetina mas elevadas, pero no se observaron aumentos en la masa de hemoglobina (Ashenden et al, 1999a, b;. Hahn et al, 2001; Dehnert et al., 2002).
Los resultados anteriores sugieren que HiHi y HiLo pueden inducir el efecto de aclimatación y aumentar el volumen total de glóbulos rojos/masa de hemoglobina si se cumplen ciertos requisitos previos (Figura 3). En los estudios HiHi que observaron aumento en el volumen total de glóbulos rojos, los atletas permanecieron 24 horas al día durante 4 semanas a una altura de aproximadamente 2500 m. En los estudios HiLo en altitud natural, el tiempo de exposición diaria a una altitud de 2500 m fue 18-20 h y la duración total fue de 3,5-4 semanas (Levine y Stray Gundersen, 1997; Wehrlin et al, 2003). En los estudios HiLo realizados en los países nórdicos utilizando una casa de altitud situada en el nivel del mar, con una hipoxia normobárica equivalente a una altura de aproximadamente 2500 m, la exposición diaria a la hipoxia fue 12-18 horas y el periodo HiLo fue de 3-4 semanas (Laitinen et al, 1995 ; Piehl-Aulin, 1999; Rusko et al, 1999). Además, en los estudios Finlandeses (Laitinen et al, 1995; Rusko et al, 1999), los atletas pasaron las horas diurnas en condiciones de hipoxia en una casa de altitud, entre las sesiones de entrenamiento a nivel del mar, y también realizaron algunas sesiones de entrenamiento de baja intensidad por semana en hipoxia, lo que podrían haber influido en sus respuestas de eritropoyetina y de volumen total de glóbulos rojos. El fracaso de los períodos de exposición cortos y tiempos de exposición diaria de corta duración para aumentar el volumen de glóbulos rojos totales/ masa de la hemoglobina (Ashenden et al, 1999a, b; Hahnetal, 2001; Dehnertetal, 2002) sugiere que el tiempo mínimo para alcanzar un efecto de aclimatación debe ser > 12 h/día a una altitud > 2000 m durante por lo menos 3 semanas (Figura 3). Las diferencias individuales observadas en el volumen total de glóbulos rojos/masa de hemoglobina pueden deberse a que los participantes podrían haber tenido deficiencia de hierro antes del período HiHi o HiLo por lo que no pudieron incrementar el volumen de glóbulos rojos (Hannon et al, 1969; Stray-Gundersen et al, 1992).
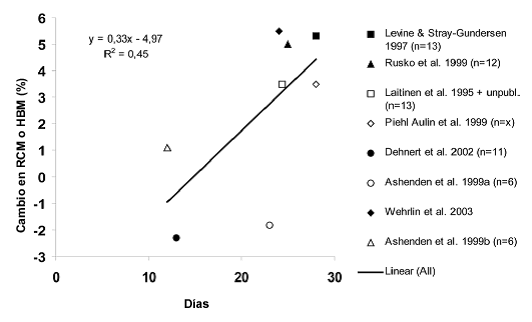
Figura 3. Las variaciones porcentuales (%) en el volumen medio total de glóbulos rojos (RCM) o masa de hemoglobina (HBM) durante HiLo en relación a la duración del período de HiLo.
PROBLEMAS CLÍNICOS RELACIONADOS CON EL ENTRENAMIENTO EN ALTITUD
Durante el entrenamiento en altura, se pueden presentar diferentes problemas médicos, tales como el mal de altura, edema cerebral o pulmonar asociados a la elevada altura. La frecuencia y gravedad de estos problemas dependen de la altitud, velocidad de ascenso y del grado de susceptibilidad individual. Los síntomas del mal de altura agudo son mas graves cuanto mayor sea la altitud por encima de 2500 m (Bartsch y Roach, 2001; Schneider et al, 2002) y estos síntomas también pueden producirse durante los períodos HiLo en una casa de altitud (HK Rusko, datos inéditos). Aunque no todos los individuos que realizan ascensos padecen estos síntomas, la hipoxia puede provocar otros efectos perjudiciales mientras se permanece en condiciones de hipoxia y algunos síntomas pueden persistir durante cierto tiempo después del regreso al nivel del mar. Por otra parte, la aclimatación previa incluso a altitudes más bajas, puede disminuir los síntomas (Bartsch y Roach, 2001). El HiLo ha sido exitoso como estrategia de aclimatación previa a HiHi, porque, por ejemplo, produce aumento en la ventilación y en la presión parcial de oxígeno en la sangre arterial en reposo y reduce el dióxido de carbono expiratorio final y la gravedad del mal de altura agudo (Beidleman et al., 2003).
La tasa metabólica cerebral global para el oxígeno y para el suministro de oxígeno al cerebro se mantendría durante la hipoxia aguda en individuos en reposo y sin mal de altura, a través de mecanismos de compensación entre los que se incluyen un aumento del flujo sanguíneo cerebral y un aumento en la concentración de hemoglobina (Roach y Hackett, 2001). Por el contrario, estudios recientes indican que durante el ejercicio en hipoxia aguda (después de 4 días a 3450 m), la saturación de oxígeno cerebral medida con espectroscopia de infrarrojo cercano puede disminuir 5-25%, y la mayor reducción se produciría en los valores superiores a 85% de la frecuencia cardíaca máxima (Bradwell et al., 1999). Por lo tanto, parecería que si la hipoxia es lo suficientemente grave, los humanos podrían experimentar una sustancial desaturación de oxígeno sistémica y cerebral durante el ejercicio en altura, y quienes presenten la desaturación cerebral más severa serán los que desarrollarán el mal de altura y edema cerebral mas severos (Roach y Hackett, 2001). Es necesario realizar estudios futuros con atletas que presentan hipoxemia arterial durante el ejercicio máximo a nivel del mar, en altitudes que se utilizan principalmente para el entrenamiento deportivo.
Aunque los mayores efectos de la hipoxia sobre la función cerebral se han observado en altitudes elevadas, incluso niveles moderados de hipoxia (>1500 m) pueden alterar la función cerebral. Se han observado síntomas como alteraciones visuales y un rendimiento mas lento, especialmente en las pruebas más complejas de la función cognitiva y motora (Hornbein, 2001). Durante la hipoxia sostenida en condiciones de altitud cada vez mayor, se han observado notables alteraciones en las pruebas neuro-psicométricas, junto con cambios en el comportamiento, estado de ánimo e incluso en la función neurológica (Hornbein, 2001). Del mismo modo, la desaturación cerebral puede afectar a diferentes sistemas críticos para el rendimiento físico. Durante la aclimatación a la hipoxia crónica, el aumento de la microvasculatura cerebral que se ha observado en estudios con animales (Harik et al., 1996) podría mejorar potencialmente la oxigenación del tejido cerebral tanto en altura como después del retorno al nivel del mar (al menos de forma transitoria), lo que reduciría el efecto limitante del sistema nervioso central sobre el rendimiento físico.
Otro mecanismo relacionado con la exposición a la altitud es la respuesta ventilatoria hipóxica. La caída en la saturación de oxígeno de la hemoglobina arterial que se produce durante el ejercicio en hipoxia, está inversamente relacionada con la respuesta ventilatoria hipóxica por lo que los sujetos que tienen una respuesta mayor pueden tener mejores rendimientos a gran altura (Ward et al, 1999, pp 69-97; Schoene, 2001).Sin embargo, la hipocapnia creciente provocada por la hiperventilación podría potencialmente disminuir la circulación cerebral (Bradwell et al., 1999; Hornbein, 2001). El resultado podría ser que, mientras se hace ejercicio en altitud, los músculos reciben más oxígeno, pero el cerebro recibe menos oxígeno, lo que puede provocar efectos secundarios negativos cuando se regresa al nivel del mar (Hornbein, 2001). Además se sabe que los atletas de resistencia tienen una quimiosensibilidad débil para el control de la ventilación en comparación con sujetos que no son atletas o que son escaladores experimentados (Schoene, 1982). Es interesante destacar que los atletas de resistencia pueden experimentar cambios en el sistema de control respiratorio y potenciar su respuesta ventilatoria hipóxica durante el período HiLo (Mattila y Rusko, 1996; Townsend et al., 2002). Esto posiblemente se debe a un aumento de la activación simpática y/o a un cambio en el punto de ajuste de la presión parcial de dióxido de carbono (PCO2) del mecanismo de control respiratorio (Townsend et al., 2002), o, como recientemente se ha hipotetizado, podría indicar un efecto primario de control respiratorio a largo plazo de la PO2 (Robbins, 2001).
Un factor que podría ocasionar problemas durante el entrenamiento en altura y disminuir el VO2max a nivel del mar después de entrenamiento en la altura es el estrés inducido por hipoxia y los consiguientes síntomas de sobreentrenamiento (Rusko, 1996; Gore et al., 1998). La activación simpática se incrementa en la altura, la cual junto con el aumento en el metabolismo glucolítico durante el ejercicio, incrementan el estrés general del entrenamiento. Se ha observado un aumento significativo en el cortisol sérico en reposo (Vasankari et al, 1993; Wilber et al, 2000) y menores concentraciones séricas de testosterona (Gore et al, 1998) después de entrenamiento en altura en atletas de resistencia de elite, lo que también podría disminuir la eritropoyesis de la médula ósea, especialmente en la etapa inicial del entrenamiento en altura (ver Berglund, 1992). La hipoxia también disminuye la respuesta inmune, específicamente a través de la supresión de la inmunidad mediada por linfocitos T (Meehan et al., 2001), y se ha observado un aumento de 50 a 100% en la frecuencia de las infecciones de las vías respiratorias superiores e infecciones gastrointestinales durante o inmediatamente después de estancias en altitud (Bailey et al, 1998; Gore et al, 1998). La posibilidad de daño oxidativo mediado por radicales libres también aumenta en condiciones de altitud (Vasankari et al., 1997).
ACLIMATACIÓN A LA ALTURA Y RENDIMIENTO DE RESISTENCIA
Varios estudios han demostrado que HiHi puede mejorar el VO2max y el rendimiento en altitud moderada (>1500 m). Durante la aclimatación, el VO2max y el rendimiento en altura comienzan a aumentar lentamente, pero el VO2max no alcanza el valor que tiene en el nivel del mar hasta después de 4 semanas de HiHi (Saltin, 1967; Faulkner et al, 1968; Adams et al., 1975, para más referencias, véase Hahn y Gore, 2001). Como pudimos observar anteriormente en esta revisión, el VO2max de los atletas de resistencia es menor a baja altura, lo que sugiere que la aclimatación podría ser beneficiosa para los rendimientos en carreras que se realizan a baja altura. Sin embargo, no hay estudios que demuestren que HiHi o HiLo podrían mejorar el rendimiento o el VO2max en altitudes bajas (500-1500 m). La literatura científica sobre HiHi para mejorar el rendimiento a nivel del mar es confusa. Aunque algunos estudios han observado mejoras (por ejemplo, Daniels y Oldridge, 1970; Mellerowicz et al, 1970), la mayoría de los estudios controlados realizados con atletas de élite no han observado un efecto positivo sobre el VO2max correspondiente al nivel del mar, el umbral de lactato, la economía de carrera o el rendimiento después de 2-4 semanas de HiHi, en comparación con un entrenamiento similar a nivel del mar, y en todos los estudios se ha observado una considerable variación individual en la respuesta al entrenamiento en altura (Svedenhag et al, 1991; Jensen et al, 1993; Rusko et al, 1996; Levine y Stray-Gundersen, 1997; Bailey et al, 1998). Los experimentos con animales también indican que, a pesar de la misma intensidad de entrenamiento absoluta en hipoxia como en normoxia, HiHi durante 10 semanas no produce ventajas significativas por encima de lo que se observa al vivir y entrenar en normoxia: el VO2max y el gasto cardíaco máximo en normoxia aumentaron de manera similar con ambos regímenes de entrenamiento (Henderson et al, 2001).
Durante HiLo, es posible lograr beneficios tanto de la aclimatación a la altura como de los estímulos de entrenamiento en normoxia (Levine y Stray-Gundersen, 1997). Varios estudios han demostrado que HiLo puede aumentar el VO2max correspondiente al nivel del mar y mejorar el rendimiento de resistencia a nivel del mar (Levine et al, 1991; Mattila y Rusko, 1996; Levine y Stray-Gundersen, 1997; Chapman et al, 1998; Piehl-Aulin , 1999; Rusko et al, 1999; Stray-Gundersen et al, 2001; Roberts et al., 2003; Wehrlin et al, 2003). En un estudio bien controlado, Levine y Stray-Gundersen (1997) demostraron que, después de 4 semanas de entrenamiento previo a nivel del mar, tanto HiLo como HiHi aumentaron VO2max a nivel del mar en proporción directa al aumento en el volumen de glóbulos rojos. Además, HiLo provocó mejoras en el umbral ventilatorio y en la velocidad en el VO2max a nivel del mar y por lo tanto en el rendimiento en carreras de pista de 5000 m que no había mejorado con HiHi o LoLo (Figura 4). La importancia de mantener una alta velocidad de entrenamiento, gasto cardíaco y flujo de oxígeno principalmente durante el entrenamiento intervalado se confirmó en otro estudio, en el cual los atletas de élite realizaron todos los ejercicios de entrenamiento de alta intensidad a baja altitud (1250 m), mientras que todo el entrenamiento inicial y de recuperación fueron realizados a una altitud de 2500 m (HiHiLo; Stray-Gundersen et al, 2001). HiHiLo indujo mejoras casi idénticas a las producidas por HiLo en el rendimiento de los atletas que hicieron todos sus ejercicios de entrenamiento a baja altura: después de 4 semanas de HiHiLo, la mejora en el VO2max a nivel del mar fue de 3% y fue acompañada por un aumento significativo de 1,1% (5,8 s ) en el rendimiento de carrera de 3000 metros a nivel del mar (Stray-Gundersen et al, 2001). En el estudio de Wehrlin et al. (2003), los atletas de resistencia de elite vivieron 24 días a aprox.2500 m, realizaron entrenamiento de base suave dos veces a la semana en una altura de ~ 1800 m, y entrenamiento intervalado dos veces a la semana en una altura de ~ 1000 m; este calendario HiHiLo provocó mejoras significativas del 4,5% en el VO2max y de 18 s en el tiempo de carrera de 5000 m.
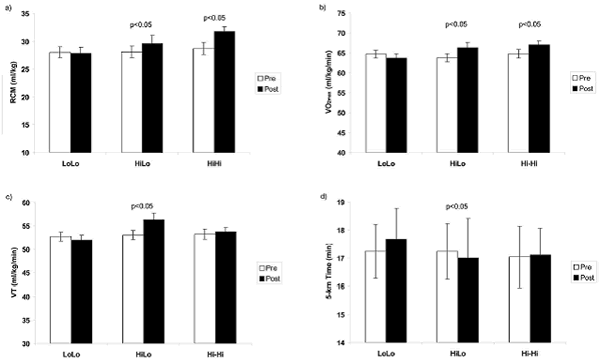
Figura 4. Comparación de los cambios observados en (a) volumen total de glóbulos rojos (RCM), (b) consumo de oxígeno máximo (VO2max), (c) umbral ventilatorio (VT) y (d) rendimiento en pruebas contrarreloj de 5000 m después de realizar entrenamiento a nivel del mar (LoLo), después de vivir alto y entrenar bajo (Hi-Lo) y después de realizar entrenamiento en altura (HiHi). Los resultados indican que tanto HiHi como HiLo aumentaron el volumen total de glóbulos rojos y el VO2max, pero sólo HiLo mejoró el umbral ventilatorio y el tiempo de pruebas contrarreloj de 5000 m. tiempo. □= Pre Entrenamiento; ■= Post entrenamiento. Modificado de Levine y Stray-Gundersen (1997).
Los efectos de Hilo y HiHiLo son igualmente individuales como los de HiHi. Los respondedores (presentaron un aumento mayor que el promedio en el rendimiento de carrera a nivel del mar) tuvieron una respuesta de eritropoyetina más marcada en altura que los no respondedores y además, los respondedores presentaron un aumento significativo en el volumen total de glóbulos rojos y en VO2max que no se observó en los no respondedores (Chapman et al, 1998). Por otra parte, los respondedores fueron más capaces que los no respondedores de mantener velocidades de entrenamiento y flujo de oxígeno normales.
Los estudios HiLo de atletas de resistencia de elite que utilizaron una casa de altitud situada a nivel del mar también observaron un aumento significativo en VO2max después de 25-28 días. Rusko et al. (1999) observaron un aumento significativo (~ 5%) en el rendimiento máximo en cinta rodante después de 25 días de HiLo y Piehl-Aulin (1999) observó un aumento significativo (~ 3%) después de 21-28 días de HiLo. También se observó una mejora de aproximadamente 4% en el rendimiento de ciclismo en ciclistas de élite después de 11 días de HiLo realizado en una casa de altitud que simulaba una altitud de 2500-3000 m (Mattila y Rusko, 1996). En contraste, no se observaron cambios significativos en el VO2max de nivel del mar ni en el rendimiento en otros estudios HiLo (Ashenden et al, 1999a, b, 2000; Hahn et al, 2001; Dehnert et al, 2002). Tal como señalamos anteriormente, una posible explicación de los resultados contradictorios es la duración mínima total de exposición a la hipoxia y la ausencia de un aumento en el volumen total de glóbulos rojos/masa de hemoglobina (Figuras 3 y 5). Los estudios que han observado un aumento significativo en el VO2max a nivel del mar y en el rendimiento después de HiLo también han observado un aumento significativo o casi significativo en el volumen total de glóbulos rojos/masa de hemoglobina (Levine y Stray-Gundersen, 1997; Piehl-Aulin, 1999; Rusko et al, 1999; Wehrlin et al, 2003), y los cambios en VO2max y en el volumen total de glóbulos rojos/masa de hemoglobina se correlacionan entre si (Levine y Stray-Gundersen, 1997; Wehrlin et al, 2003).
En otro estudio HiLo, los atletas que habían vivido y entrenado por un año en altura, presentaron una mejora significativa en el rendimiento de resistencia después de un período de entrenamiento simultáneo de 6 semanas HiLo mientras respiraban 70% de oxígeno durante cuatro sesiones semanales de entrenamiento en altitud, lo que permitió una mayor intensidad de entrenamiento (Chick et al, 1993). Del mismo modo, respirar gas hiperóxico durante tres sesiones de alta intensidad por semana durante 3 semanas, a una altitud de 1840 m permitió una mayor intensidad absoluta de entrenamiento (Wilber et al, 2003) y provocó una mejora en el estado estacionario máximo de lactato y en el tiempo necesario para completar un test de rendimiento de 120 kJ (Morris et al, 2000). En un experimento con animales también se observó un mayor rendimiento de carrera, menor frecuencia cardíaca a velocidad de carrera constante y mayor masa del ventrículo izquierdo y del tríceps sural en el grupo HiLo en comparación con el grupo LoLo (Miyazaki y Sakai, 2000).
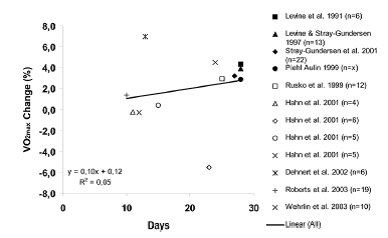
Figura 5. Cambio porcentual (%) en el consumo de oxígeno máximo medio (VO2max) durante HiLo en relación a la cantidad de días del período de HiLo.
Piehl Aulin (1999) observó una mejora significativa en el VO2max a nivel del mar durante HiLo (día 21), así como inmediatamente después y 1 semana después del período de 28 días de HiLo. Levine y Stray-Gundersen (1997) observaron mejoras significativas tanto en el VO2max a nivel del mar como en el rendimiento 3, 7, 14 y 21 días después de HiLo. En contraste, Rusko et al. (1999) observaron mejoras en el rendimiento, VO2max, en el rendimiento máximo en cinta rodante y en el rendimiento de ciclismo (Mattila y Rusko, 1996) 1 semana después del período HiLo pero no inmediatamente (unas horas) después de la última noche en hipoxia (Rusko et al, 1999). En varios estudios realizados en Australia, se realizaron post-tests “dentro de las 36 horas” después del período de HiLo y no se observaron cambios significativos (Ashenden et al, 1999a, b, 2000; Hahn et al, 2001).
La mejora el rendimiento de resistencia a nivel del mar podría también deberse a un aumento en la capacidad aeróbica de los músculos o a la mejora de la economía después de HiHi e HiLo. Quienes viven a gran altitud tienen una mayor concentración de mioglobina muscular y una mayor capacidad respiratoria muscular que quienes residen al nivel del mar (Reynafarje, 1962). Sin embargo, en los atletas de élite se ha demostrado que ni HiHi ni HiLo aumentan la oxidación ni la capacidad glucolítica de los músculos (Rahkila y Rusko, 1982; Saltin et al, 1995a, b; Stray-Gundersen et al., 1999). Los resultados de los estudios sobre el efecto que tiene el entrenamiento en altura en la economía son contradictorios (Svedenhag et al, 1991; Saltin et al, 1995a; Levine y Stray-Gundersen, 1997; Green et al, 2000;.. Gore et al, 2001).
ACLIMATACIÓN A LA ALTURA Y RENDIMIENTO ANAERÓBICO
Debido a la disminución de la resistencia del aire, se registraron nuevas marcas mundiales en todos los eventos de esprint en los Juegos Olímpicos de México en 1968. Karvonen et al. (1990) demostraron que las velocidades máximas de esprint aumentaron durante 2 semanas de entrenamiento en altura a 1860 m, probablemente debido a la disminución de la resistencia del aire durante el entrenamiento de esprint. Gale y Nagle (1971) demostraron que el entrenamiento de esprint de ratas realizado a aprox. 2300 m de altitud durante 5 semanas a la misma velocidad que en normoxia aumentó las reservas de fosfocreatina en el músculo gastrocnemio-plantar.
Durante la exposición a la altitud, los rendimientos supramáximos dependientes de la producción de energía gucolítica disminuirían debido a la pérdida de bicarbonato y a la disminución en el volumen de plasma (Hahn y Gore, 2001). De hecho, al realizar 10 series x 6-s de ejercicio intervalado supramáximo, se vio afectada la capacidad de mantener una elevada producción de potencia durante la última serie, el VO2max fue menor y la acumulación de lactato fue mayor en hipoxia (aprox.3000 m) que en normoxia (Balsom y col., 1994). Sin embargo, la hipoxia aguda, no afectó el rendimiento en un único test de Wingate de 30-45 s, a pesar de que disminuyó el VO2max, aumentó el lactato muscular y disminuyó el glucógeno después del ejercicio en hipoxia en comparación con la situación de normoxia (McLellan et al, 1990). Se ha demostrado que la ingesta o infusión de bicarbonato de sodio aumenta la capacidad buffer de la sangre en altura, disminuye la desaturación arterial y mejora el rendimiento y el entrenamiento intervalado en altura (Hausswirth et al, 1995 Nielsen et al, 2002), lo que indica que en condiciones de altitud disminuye el potencial para el entrenamiento y la producción de energía glucolítica y no se producen aumentos compensatorios en la capacidad buffer. Otras mediciones para mejorar el rendimiento en las carreras y la intensidad del entrenamiento en altitud podría ser la ingesta de cafeína (Berglund y Hemmingsson, 1982) y una dieta alta en carbohidratos (Hansen et al, 1972).
Se ha observado que el HiHi crónico durante 3-4 semanas no mejora la capacidad de rendimiento anaeróbico supramaximo al nivel del mar (Rusko et al, 1996; Levine y Stray-Gundersen, 1997; Bailey et al., 1998). Sin embargo, se ha demostrado que el entrenamiento en una altura por encima de 2000 m durante 2 semanas aumenta el tiempo de carrera hasta el agotamiento (rango de 240 a 380 s) durante la medición del VO2max a nivel del mar sin aumento en VO2max, y aumenta el déficit de oxígeno acumulado máximo y la capacidad buffer muscular después del retorno al nivel del mar (Mizuno et al, 1990; Svedenhag et al, 1991; Saltin et al, 1995b). Martino et al. (1996) también demostraron que el rendimiento anaeróbico a nivel del mar puede mejorar después de realizar entrenamiento esprint de natación en altura.
También se ha demostrado que el entrenamiento HiLo durante 2 semanas con 14-18 h de exposición diaria a hipoxia normobarica en una casa de altitud (15,8% de O2 en el aire inspirado, aprox. 2500 m) y el entrenamiento normal de sprint a nivel del mar pueden mejorar el tiempo de carrera de 400 m a nivel del mar de velocistas de élite, y disminuir la concentración de lactato sanguíneo durante esprints submáximos en una cinta rodante (Nummela y Rusko, 2000). Gore et al. (2001) demostraron que 23 días de HiLo, en una casa de altitud, con una altitud equivalente a 3000 m, aumentaron la capacidad buffer muscular. En un reciente estudio australiano, ninguna de las duraciones utilizadas de HiLo de 5, 10 o 15 días, 8-10 h por noche, produjo cambios significativos en VO2max, déficit de oxígeno máximo acumulado ni en la producción de potencia media máxima en 4 min (MMPO4min), pero cuando se utilizaron los datos combinados de los tres períodos evaluados, se observó un aumento significativo en el déficit de oxígeno acumulado máximo y en MMPO4min (Roberts et al, 2003). Aunque Levine y Stray-Gundersen (1997) no observaron una mejora en déficit de oxígeno acumulado máximo y Stray-Gundersen et al. (1999) observaron una disminución de la capacidad buffer muscular después de 4 semanas de HiLo, en atletas que realizaban entrenamiento de resistencia aeróbica, parecería que HiHi e HiLo con entrenamiento de tipo sprint pueden mejorar el rendimiento anaeróbico subsiguiente realizado al nivel del mar.
EFECTOS DEL ENTRENAMIENTO HIPÓXICO
Los estudios donde se experimentó hipoxia sólo durante las sesiones de entrenamiento no han observado efectos de aclimatación (ej. Roskam et al, 1969; Terrados et al, 1988; Geiser et al, 2001; Hendriksen y Meeuwsen, 2003), a pesar de que se observó un aumento en la eritropoyetina después del ejercicio en condiciones de hipoxia (Schmidt et al, 1991).
Se ha demostrado que el entrenamiento en hipoxia aumenta el VO2max y el rendimiento submáximo y máximo al nivel del mar, pero los efectos han sido similares a los del entrenamiento en normoxia (Roskam et al, 1969; Davies y Sargeant, 1974; Terrados et al, 1988; Burtscher et al., 1996). En un reciente estudio con diseño cruzado, de entrenamiento hipóxico de intensidad moderada (60-70% VO2max) durante 10 días se observó un aumente en la producción de potencia máxima aeróbica y anaeróbica en normoxia después de 2-9 días de haber finalizado el entrenamiento hipóxico. Sin embargo, el aumento de la producción de potencia aeróbica máxima no fue significativamente mayor que después del entrenamiento control en normoxia, y el VO2max no cambió significativamente (Hendriksen y Meeuwsen, 2003), a pesar de que se observaron algunos cambios significativos durante la primera mitad del estudio cruzado (Meeuwsen et al, 2001). Del mismo modo, en atletas entrenados, el entrenamiento en hipoxia tres veces por semana durante 6 semanas, no provocó un mejor rendimiento ni en condiciones de normoxia ni en hipoxia (Ventura et al., 2003). Según Geiser et al. (2001), no se observaron diferencias entre los efectos del entrenamiento en hipoxia y en normoxia en individuos no entrenados cuando se midió el VO2max en normoxia, pero el entrenamiento en hipóxia provocó un aumento en el VO2max en hipoxia mayor que el del entrenamiento en normoxia, y el entrenamiento de alta intensidad en hipoxia fue el estímulo más eficaz para aumentar la capacidad oxidativa muscular. Terrados et al. (1988) también demostraron que el entrenamiento en hipoxia provocó un aumento en la capacidad de trabajo de los atletas en hipoxia mayor al del entrenamiento en normoxia a nivel del mar; el incremento relacionó con una disminución en la concentración de lactato sanguíneo durante el ejercicio, con un incremento en la capilarización y con una disminución de la capacidad glucolítica. De hecho, cuando la intensidad absoluta del entrenamiento en hipoxia era igual a la del entrenamiento en normoxia (por ejemplo, ejercicios de piernas individuales en hipoxia y normoxia), se observó un mejor rendimiento a nivel del mar y un aumento en la mioglobina y en la capacidad oxidativa en los músculos humanos (Terrados et al, 1990; Melissa et al, 1997; Hoppeler y Vogt, 2001).
Los resultados anteriores sugieren que el entrenamiento en hipoxia induce cambios similares a los del entrenamiento en normoxia para mejorar el rendimiento a nivel del mar, pero el entrenamiento en hipoxia puede ser más beneficioso que el entrenamiento a nivel del mar para mejorar el rendimiento en altura.
HIPOXIA INTERMITENTE A CORTO PLAZO EN REPOSO
La hipoxia intermitente a corto plazo en reposo (IHR) fue desarrollada originalmente en la antigua Unión Soviética y consiste en respirar aire hipóxico (9-11%) a través de una máscara o boquilla durante períodos repetidos de 5-7 minutos, interrumpidos por períodos iguales de recuperación en normoxia, durante un total de 1-3 h en una o dos sesiones al día (Bernardi, 2001; Serebrovskaya, 2002). Como prueba de que la hipoxia intermitente a corto plazo en reposo podría mejorar la aclimatación a la altura, Tkatchouk et al. (1999) observaron un aumento en la presión parcial de oxígeno en la sangre arterial (de 54±1 a 62±2 mmHg), P(a-v)O2 (de 22±3 a 31±1 mmHg) y en la saturación de oxígeno de la hemoglobina arterial (de 87 a 91%) durante el ascenso agudo a 3000 m en una cámara de altitud en comparación con los valores previos a IHR. Según lo observado por Serebrovskaya (2002), quien revisó los estudios llevados a cabo en la antigua Unión Soviética y en la Comunidad de Estados Independientes, la hipoxia intermitente en descanso indujo “una mayor sensibilidad ventilatoria a la hipoxia, así como también otros cambios fisiológicos relacionados con la hipoxia, tales como aumentos en la hematopoyesis, capacidad de ventilación alveolar y difusión pulmonar, y alteraciones en el sistema nervioso autónomo. Los efectos positivos de la hipoxia intermitente a corto plazo en reposo para mejorar el rendimiento a nivel del mar han sido observados en estudios realizados con nadadores competitivos evaluados frente a un grupo control (Bulgakova et al, 1999).
También hay estudios realizados en Occidente que observaron efectos positivos de aclimatación gracias a la hipoxia intermitente a corto plazo en reposo lo que sugiere que la exposición de 3-5 horas al día durante 2-4 semanas a RSI severa de corta duración, podría producir incrementos en el rendimiento, la concentración de hemoglobina, el hematocrito y los reticulocitos (Hellemans, 1999; Rodríguez et al, 1999, 2003; Casas et al, 2000). Estos resultados deben ser confirmados porque los estudios no contemplaron la presencia de un grupo control, no realizaron medición del volumen total de glóbulos rojos/masa de hemoglobina y porque algunos estudios incluyeron entrenamiento en hipoxia. Del mismo modo, en un estudio controlado, doble ciego de 4 semanas, 5 días después de un programa RSI-respiratoria (5 sesiones por semana, 9-11% O2), el grupo experimental (n = 14) formado por estudiantes de deportes presentó menores valores de frecuencia cardiaca, ventilación por minuto y doble producto (frecuencia cardiaca x presión arterial sistólica) en comparación con el grupo placebo (w=14) con una carga de trabajo de ciclismo constante de 150 w (Burtscher et al, 1999). En contraste, en un estudio Australiano (Clark et al, 1999), ocho remeros de élite fueron agrupados de a dos en función de sus VO2max y del rendimiento. El grupo tratamiento inhaló 12,2% de O2 y el grupo control 20,9% de O2 en una relación de 5 min tratamiento/5 min recuperación durante un total de 90 minutos por día durante 14 días. Durante la hipoxia intermitente en reposo, mediante oximetría de pulso se pudo observar una disminución en la saturación de oxígeno de la hemoglobina arterial (P<0,05) y un aumento de la frecuencia cardíaca (P= 0,06) en el grupo experimental en comparación con el grupo control. Sin embargo, no se observó ningún cambio en los valores en reposo del volumen total de glóbulos rojos, concentración de hemoglobina, hematocrito y reticulocitos entre los valores pre y post-tratamiento para ninguno de los grupos. Durante la prueba en ergómetro de remo, no se observaron diferencias en la frecuencia cardíaca, VO2 submáximo, VO2max, capacidad de ejercicio, concentración de lactato y tasa de intercambio respiratorio.
Los resultados anteriores sugieren que la hipoxia intermitente a corto plazo en reposo no se traduce en un efecto hematológico de aclimatación y es dudoso que pueda mejorar los estímulos de entrenamiento para mejorar el rendimiento a nivel del mar sin hacer ejercicio en hipoxia. Sin embargo, podría mejorar la adaptación a la posterior exposición y entrenamiento en altura.
EFECTOS DE ARRASTRE DE “VIVIR ALTO” PARA EL POSTERIOR “ENTRENAR BAJO”
Algunos efectos de vivir alto continúan ejerciendo su influencia en el entrenamiento subsiguiente en normoxia, entre los cuales podemos mencionar: disminución en el volumen plasmático, mayor concentración de hemoglobina y aumento en el volumen total de glóbulos rojos/masa de hemoglobina (Levine y Stray-Gundersen, 1997); aumento en la activación simpática y parasimpática (Boushel et al., 2001); y aumento de la respuesta ventilatoria hipóxica y la respuesta ventilatoria al ejercicio durante y después de HiLo (Mattila y Rusko, 1996; Ward et al, 1999, pp 69-97; Townsend et al, 2002). Una mejora en la respuesta ventilatoria hipóxica es potencialmente una adaptación positiva para el entrenamiento al nivel del mar debido a que un aumento en la ventilación aumenta la presión alveolar de oxígeno y mejora la oxigenación arterial. En consecuencia, la saturación de oxígeno de la hemoglobina arterial determinada mediante oximetría de pulso y el contenido de oxígeno arterial se incrementan en normoxia durante HiLo y después HiHi y HiLo, y la saturación de oxígeno de la hemoglobina arterial determinada mediante oximetría de pulso durante el ejercicio en normoxia, puede aumentar y acercarse a los valores previos observados a nivel del mar (HK Rusko, datos inéditos). El aumento en 2,3DPG inducido por vivir en hipoxia también podría aumentar la extracción de oxígeno durante el entrenamiento posterior en normoxia (Mairbäurl, 1994). Por lo tanto, aumenta la posibilidad de aumentar la intensidad de entrenamiento durante el entrenamiento posterior en normoxia durante y después de HiLo. De hecho, se ha observado una disminución en la concentración de lactato sanguíneo durante el ejercicio submáximo al regresar al nivel del mar un día después de un campamento de entrenamiento en altura, en comparación con el entrenamiento pre-altitud; y la disminución de la concentración de lactato sanguíneo fue mayor cuando mayores fueron los valores de la concentración de hemoglobina y del hematocrito (Ingjer y Myhre, 1992).
CONCLUSIONES
Los beneficios para los atletas de vivir alto y entrenar alto (HiHi) para mejorar el rendimiento en altitud son claros, pero los estudios controlados para un mejor rendimiento al nivel del mar son controvertidos. Las razones de la ausencia de un efecto positivo de HiHi son las siguientes: (1) en los atletas de élite, el efecto de aclimatación puede haber sido insuficiente para estimular un aumento en el volumen total de glóbulos rojos/masa de hemoglobina, debido a que la altitud fue insuficiente (<2000-2200 m) y/o a un periodo de tiempo en altura inadecuado (<3-4 semanas); (2) el efecto del entrenamiento en altitud podría haber sido afectado por la insuficiencia de estímulos de entrenamiento para mejorar la función de los sistemas neuromuscular y cardiovascular; y (3) un aumento en el estrés con posibles síntomas de sobreentrenamiento y una mayor frecuencia de infecciones. Al comparar HiHi con HiLo, está claro que ambos entrenamientos provocan un efecto positivo de aclimatación y aumentan la capacidad de transporte de oxígeno de la sangre, por lo menos en los “respondedores'', siempre que se cumplen ciertos requisitos previos. El tiempo mínimo para alcanzar un efecto de aclimatación debe ser >12 h al día durante al menos 3 semanas, a una altitud o altura simulada de 2100-2500 m. La exposición a hipoxia parece tener algunos efectos de transferencia positivos para el entrenamiento posterior en normoxia durante y después de HiLo. La mayor capacidad de transporte de oxígeno de la sangre posibilita el entrenamiento en intensidades más altas después de HiHi y durante y después de HiLo en la subsiguiente normoxia, lo que aumenta los estímulos de entrenamiento y la mejora de algunos determinantes neuromusculares y cardiovasculares de la capacidad de resistencia. Los efectos de la hipoxia en el cerebro pueden influir tanto en la intensidad del entrenamiento como en las respuestas fisiológicas durante el entrenamiento en hipoxia. Por otra parte, la interrupción de la exposición a la hipóxia por entrenamiento en normoxia puede ser un factor clave para evitar o minimizar los efectos nocivos, que se sabe que ocurren en condiciones de hipoxia crónica. Los efectos del entrenamiento hipóxico y de la hipoxia severa a corto plazo en reposo aún no son claros y deben ser estudiados con mayor detalle.
Referencias
1. Adams, W.C., Bernauer, E.M., Dill, D.B. and Bomar, J.B., Jr. (1975). Effects of equivalent sea level and altitude training on VO2max and running performance. Journal of Applied Physiology, 39, 262–266.
2. Ashenden, M.J., Gore, C.J., Dobson, G.P. and Hahn, A.G. (1999a). ‘Live high, train low’ does not change the total haemoglobin mass of male endurance athletes sleeping at a simulated altitude of 3000 m for 23 nights. European Journal of Applied Physiology, 80, 479–484.
3. Ashenden, M.J., Gore, C.J., Martin, D.T., Dobson, G.P. and Hahn, A.G. (1999b). Effects of a 12-day ‘live high, train low’ camp on reticulocyte production and haemoglobin mass in elite female road cyclists. European Journal of Applied Physiology, 80, 472–478.
4. Ashenden, M.J., Gore, C.J., Dobson, G.P. et al. (2000). Simulated moderate altitude elevates serum erythropoietin but does not increase reticulocyte production in well-trained runners. European Journal of Applied Physiology, 81, 428–435.
5. Bailey, D.M. and Davies, B. (1997). Physiological implications of altitude training for endurance performance at sea level: a review. British Journal of Sports Medicine, 31, 183– 190.
6. Bailey, D.M., Davies, B., Romer, L. et al. (1998). Implications of moderate altitude training for sea-level endurance in elite distance runners. European Journal of Applied Physiology, 78, 360–368.
7. Balsom, P.D., Gaitanos, G.C., Ekblom, B. and Sjo¨din, B. (1994). Reduced oxygen availability during high intensity intermittent exercise impairs performance. Acta Physiologica Scandinavica, 152, 279–285.
8. Ba¨rtsch, P. and Roach, R. (2001). Acute mountain sickness and high-altitude cerebral edema. In: High Altitude: An Exploration of Human Adaptation (edited by T.F. Hornbein and R.B. Schoene), pp. 731–776. New York: Marcel Dekker.
9. Beidleman, B.A., Muza, S.R., Fulco, C.S. et al. (2003). Intermittent altitude exposures eliminate acute mountain sickness at 4300 m. Medicine and Science in Sports and Exercise, 35, S163.
10. Benoit, H., Busso, T., Castells, J., Geyssant, A. and Denis, C. (2003). Decrease in peak heart rate with acute hypoxia in relation to sea level. Published online on 26 July in European Journal of Applied Physiology (available at: http:// www.springerlink.com).
11. Berglund, B. (1992). High-altitude training: aspects of hematological adaptation. Sports Medicine 14, 289–303.
12. Berglund, B. and Hemmingsson, P. (1982). Effects of caffeine ingestion on exercise performance at low and high altitudes in cross-country skiers. International Journal of Sports Medicine, 3, 234–236.
13. Bernardi, L. (2001). Interval hypoxic training. In Hypoxia: From Genes to the Bedside. Proceedings of the 12th Interna-tional Hypoxia Symposium (edited by R.C. Roach, P.D. Wagner and P.H. Hackett), pp. 377–399. New York: Kluwer Academic/Plenum Publishers.
14. Bo¨ning, D., Rojas, J., Serrato, M. et al. (2001). Hemoglobin mass and peak oxygen uptake in untrained and trained residents of moderate altitude. International Journal of Sports Medicine, 22, 572–578.
15. Boushel, R., Calbet, J.A., Radegran, G. et al. (2001). Parasympathetic neural activity accounts for the lowering of exercise heart rate at high altitude. Circulation, 104, 1785–1791.
16. Bradwell, A.R., Williams, D., Beazley, M. and Imray, C.H.E. (1999). Can acute mountain sickness be induced by exercise? In Hypoxia: Into the Next Millennium. Proceedings of the 11th International Hypoxia Symposium (edited by R.C. Roach, P.D. Wagner and P.H. Hackett), pp. 368– 369. New York: Kluwer Academic/Plenum Publishers.
17. Brosnan, M.J., Martin, D.T., Hahn, A.G., Gore, C.J. and Hawley, J.A. (2000). Impaired interval exercise responses in elite female cyclists at moderate simulated altitude. Journal of Applied Physiology, 89, 1819–1824.
18. Bulbulian, R., Wilcox, A.R. and Darabos, B.L. (1986). Anaerobic contribution to distance running performance of trained cross-country athletes. Medicine and Science in Sports and Exercise, 21, 107–113.
19. Bulgakova, N.J., Kovalev, N.V. and Volkov, N.I. (1999). Interval hypoxia training enhances effects of physical loads in swimming. In Biomechanics and Medicine in Swimming VIII. Proceedings of the VIII International Symposium on Biomechanics and Medicine in Swimming (edited by K.L. Keskinen, P.V. Komi and A.P. Hollander), pp. 413–415. Jyvaskyla¨: University of Jyvaskyla¨, Department of Biology of Physical Activity.
20. Burtscher, M., Nachbauer, W., Baumgartl, M. and Philadelphy, M. (1996). Benefits of training at moderate altitude versus sea level training in amateur runners. European Journal of Applied Physiology, 74, 558–563.
21. Burtscher, M., Tsvetkova, A.M., Tkatchouk, E.N. et al. (1999). Beneficial effects of short term hypoxia. In Hypoxia: Into the Next Millennium. Proceedings of the 11th International Hypoxia Symposium (edited by R.C. Roach, P.D. Wagner and P.H. Hackett), pp. 371–372. New York: Kluwer Academic/Plenum Publishers.
22. Carter, J., Jones, D., Mann, C. and Jeukendrup, A. (2003). The effects of glucose infusion on 1 h cycle time trial performance. In Proceedings of the 8th Annual Congress of the European College of Sport Science (edited by E. Mu¨ller, H. Schwameder, G. Zallinger and V. Fastenbauer), pp. 29–30. Salzburg: Institute of Sport Science, University of Salzburg.
23. Casas, M., Casas, H., Page´s, T. et al. (2000). Intermittent hypobaric hypoxia induces altitude acclimatization and improves the lactate threshold. Aviation, Space and Environmental Medicine, 71, 125–130.
24. Chapman, R.F., Stray-Gundersen, J. and Levine, B.D. (1998). Individual variation in response to altitude training. Journal of Applied Physiology, 85, 1448–1456.
25. Chick, T.W., Stark, D.M. and Murata, G.H. (1993). Hyperoxic training increases work capacity after maximal training at moderate altitude. Chest, 104, 1759– 1762.
26. Clark, S.A., Dixon, J., Gore, C.J. and Hahn, A.G. (1999). 14 days of intermittent hypoxia does not alter haematological parameters amongst endurance trained athletes (avail-able at: http://www. ausport.gov.au/fulltext/1999/triathlon/ clark.dixon.gor.hahn.pdf).
27. Daniels, J. and Oldridge, N. (1970). The effects of alternate exposure to altitude and sea level on world-class middle-distance runners. Medicine and Science in Sports and Exercise, 2, 107–112.
28. Davies, C.T.M. and Sargeant, A.J. (1974). Effects of hypoxic training on normoxic maximal aerobic power. European Journal of Applied Physiology, 33, 227–236.
29. Dehnert, C., Huütler, M., Liu, Y. et al. (2002). Erythropoiesis and performance after two weeks of living high and training low in well trained athletes. International Journal of Sports Medicine, 23, 561–566.
30. Dempsey, J.A. and Wagner, P.D. (1999). Exercise-induced arterial hypoxemia. Journal of Applied Physiology, 87, 1997– 2006.
31. Dempsey, J.A., Hanson, P.G. and Henderson, K.S. (1984). Exercise-induced arterial hypoxaemia in healthy human subjects at sea level. Journal of Physiology, 355, 161–175.
32. Di Lazzaro, V., Oliviero, A., Tonali, P.A. et al. (2003). Direct demonstration of reduction of the output of the human motor cortex induced by a fatiguing muscle contraction. Experimental Brain Research, 149, 535–538.
33. Eckhardt, K.-U., Boutellier, U., Kurtz, A., Schopen, M., Koller, E.A., et al. (1989). Rate of erythropoietin forma-tion in response to acute hypobaric hypoxia. Journal of Applied Physiology, 66, 1785–1788.
34. Faulkner, J.A., Kollias, J., Favour, C.B., Buskirk, E.R. and Balke, B. (1968). Maximum aerobic capacity and running performance at altitude. Journal of Applied Physiology, 24, 685–691.
35. Fiori, G., Facchini, F., Ismagulov, O. et al. (2000). Lung volume, chest size, and haematological variation in low-, medium-, and high-altitude central Asian populations. American Journal of Physical Anthropology, 113, 47–59.
36. Gale, J.B. and Nagle, G. (1971). Changes in ATP and creatine phosphate storage in skeletal muscle of rats trained at 900 and 7600 feet. Nature, 232, 30.
37. Gandevia, S.C. (2001). Spinal and supraspinal factors in human muscle fatigue. Physiological Review, 81, 1725– 1789.
38. Geiser, J., Vogt, M., Billeter, R. et al. (2001). Training high– living low: changes of aerobic performance and muscle structure with training at simulated altitude. International Journal of Sports Medicine 22, 579–585.
39. Gore, C.J., Hahn, A.G., Watson, D.B. et al. (1995). VO2max & arterial O2 saturation at sea level & 610 m. Medicine and Science in Sports and Exercise, 27, S7
40. Gore, C.J., Hahn, A.G., Scroop, G.C. et al. (1996). Increased arterial desaturation in trained cyclists during maximal exercise at 580 m altitude. Journal of Applied Physiology, 80, 2204–2210.
41. Gore, C.J., Little, S.C., Hahn, A.G. et al. (1997). Reduced performance of male and female athletes at 580 m altitude. European Journal of Applied Physiology, 75, 136–143.
42. Gore, C., Craig, N., Hahn, A. et al. (1998). Altitude training at 2690 m does not increase total haemoglobin mass or sea level VO2max in world champion track cyclists. Journal of Science and Medicine in Sport, 1, 156–170.
43. Gore, C.J., Hahn, A.G., Aughey, R.J. et al. (2001). Live high:train low increases muscle buffer capacity and submaximal cycling efficiency. Acta Physiologica Scandinavica, 173, 275–286.
44. Green, H.J., Roy, B. and Grant, S. (2000). Increases in submaximal cycling efficiency mediated by altitude acclimatization. Journal of Applied Physiology, 89, 1189–1197.
45. Grover, R.F. and Bärtsch, P. (2001). Blood. In High Altitude: An Exploration of Human Adaptation (edited by T.F. Hornbein and R.B. Schoene), pp. 493–523. New York: Marcel Dekker.
46. Hahn, A.G. and Gore, C.J. (2001). The effect of altitude on cycling performance: a challenge to traditional concepts. Sports Medicine, 31, 533–557.
47. Hahn, A.G., Gore, C.J., Martin, D.T. et al. (2001). An evaluation of the concept of living at moderate altitude and training at sea level. Comparative Biochemistry and Physiology. Part A, 128, 777–789.
48. Hannon, J.P., Shields, J.L. and Harris, C.W. (1969). Effects of altitude acclimatization on blood composition of women. Journal of Applied Physiology, 26, 540–547.
49. Hansen, J.E., Hartley, H. and Hogan, R.P. (1972). Arterial oxygen increased by high-carbohydrate diet at altitude. Journal of Applied Physiology, 83, 441–445.
50. Harik, N., Harik, S.I., Kuo, N.T. et al. (1996). Time-course and reversibility of the hypoxia-induced alterations in cerebral vascularity and cerebral capillary glucose trans-porter density. Brain Research, 737, 335–338.
51. Hausswirth, C., Bigard, A.-X., Lepers, R., Berthelot, M. and Guezennec, C.Y. (1995). Sodium citrate ingestion and muscle performance in acute hypobaric hypoxia. European Journal of Applied Physiology, 71, 362–368.
52. Heinicke, K., Prommer, N., Cajigal, J. et al. (2003). Long-term exposure to intermittent hypoxia results in increased hemoglobin mass, reduced plasma volume, and elevated erythropoietin plasma levels in man. European Journal of Applied Physiology, 88, 535–543.
53. Hellemans, J. (1999). Intermittent hypoxic training: a pilot study (available at: http://www. go2altitude.com/ dr_hellemans.html).
54. Henderson, K.K., Clancy, R.L. and Gonzalez, N.C. (2001). Living and training in moderate hypoxia does not improve VO2max more than living and training in normoxia. Journal of Applied Physiology, 90, 2057–2062.
55. Hendriksen, I.J.M. and Meeuwsen, T. (2003). The effect of intermittent training in hypobaric hypoxia on sea-level exercise: a cross-over study in humans. European Journal of Applied Physiology, 88, 396–403.
56. Hoppeler, H. and Vogt, M. (2001). Hypoxia training for sea-level performance. In Hypoxia: From Genes to the Bedside. Proceedings of the 12th International Hypoxia Symposium (edited by R.C. Roach, P.D. Wagner and P.H. Hackett), pp. 61–73. New York: Kluwer Academic/Plenum Publishers.
57. Hornbein, T.F. (2001). The high-altitude brain. Journal of Experimental Biology, 204, 3129–3132.
58. Houmard, J.A., Scott, B.K., Justice, C.L. and Chenier, T.C. (1994). The effects of taper on performance in distance runners. Medicine and Science in Sports and Exercise, 26, 624–631.
59. Ingjer, F. and Myhre, K. (1992). Physiological effects of altitude training on young elite male cross-country skiers. Journal of Sports Sciences, 10, 37–47.
60. Jelkmann, W. and Hellwig-Bürgel, T. (2001). Biology of erythropoietin. In Hypoxia: From Genes to the Bedside. Proceedings of the 12th International Hypoxia Symposium (edited by R.C. Roach, P.D. Wagner and P.H. Hackett), pp. 169–187. New York: Kluwer Academic/Plenum Publishers.
61. Jensen, J.D., Jensen, L.W., Madsen, J.K. and Poulsen, L. (1995). The metabolism of erythropoietin in liver cirrhosis patients compared with healthy volunteers. European Journal of Haematology, 54, 111–116.
62. Jensen, K., Nielsen, T.S., Fiskestrand, A. et al. (1993). High-altitude training does not increase maximal oxygen uptake or work capacity at sea level in rowers. Scandinavian Journal of Medicine and Science in Sports, 3, 256–262.
63. Johnston, R.E., Quinn, T.J., Kertzer, Rand Vroman, N.B. (1997). Strength training in female distance runners: impact on running economy. Journal of Strength Conditioning Research, 11, 224–229.
64. Karvonen, J., Peltola, E., Saarela, J. and Nieminen, M. (1990). Changes in running speed, blood lactic acid concentration and hormone balance during sprint training performed at an altitude of 1860 metres. Journal of Sports Medicine and Physical Fitness, 30, 122–126.
65. Kay, D., Marino, F., Cannon, J. et al. (2001). Evidence for neuromuscular fatigue during high intensity cycling in warm, humid conditions. European Journal of Applied Physiology, 84, 115–121.
66. Kayser, B., Narici, M., Binzoni, T., Grassi, B. and Cerretelli, P. (1994). Fatigue and exhaustion in chronic hypobaric hypoxia: influence of exercising muscle mass. Journal of Applied Physiology, 76, 634–640.
67. Koistinen, P.O., Rusko, H., Irjala, K. et al. (2000). EPO, red cells, and serum transferrin receptor in continuous and intermittent hypoxia. Medicine and Science in Sports and Exercise, 32, 800–804.
68. Lahiri, S., Di Giulio, C. and Roy, A. (2002). Lessons from chronic intermittent and sustained hypoxia at high altitudes. Respiratory, Physiology and Neurobiology, 130, 223–233.
69. Laitinen, H., Alopaeus, K., Heikkinen, R. et al. (1995). Acclimatization to living in normobaric hypoxia and training in normoxia at sea level in runners. Medicine and Science in Sports and Exercise, 27, S109.
70. Levine, B.D. (2002). Intermittent hypoxic training: fact and fancy. High Altitude Medicine and Biology, 3, 177–194.
71. Levine, B.D. and Stray-Gundersen, J. (1997). ‘Living high– training low’: effect of moderate-altitude acclimatization with low altitude training on performance. Journal of Applied Physiology, 83, 102–112.
72. Levine, B.D., Stray-Gundersen, J., Duhaime, G., Snell, P.G. and Friedman, D.B. (1991). Living high–training low: the effect of altitude acclimatization/normoxic training in trained runners. Medicine and Science in Sports and Exercise, 23, S25.
73. Lundby, C., Araoz, M. and van Hall, G. (2001a). Peak heart rate decreases with increasing severity of acute hypoxia. High Altitude Medicine and Biology, 2, 369–376.
74. Lundby, C., Moeller, P., Kanstrup, I.-L. and Olsen, N.V. (2001b). Heart rate response to hypoxic exercise: role of dopamine D2-receptors and effect of oxygen supplementation. Clinical Science, 101, 377–383.
75. Mairbaürl, H. (1994). Red blood cell function in hypoxia at altitude and exercise. International Journal of Sports Medicine, 15, 51–63.
76. Martino, M., Myers, K. and Bishop, P. (1996). Effetcs of 21 days training at altitude on sea-level anaerobic performance in competitive swimmers. Medicine and Science in Sports and Exercise, 27, S5.
77. Mattila, V. and Rusko, H. (1996). Effect of living high and training low on sea level performance in cyclists. Medicine and Science in Sports and Exercise, 28, S156.
78. McLellan, T.M., Kavanagh, M.F. and Jacobs, I. (1990). The effect of hypoxia on performance during 30 s or 45 s of supramaximal exercise. European Journal of Applied Physiology, 60, 155–161.
79. Meehan, R.T., Uchakin, P.N. and Sams, C.F. (2001). High altitude and human immune responsiveness. In High Altitude: An Exploration of Human Adaptation (edited by T.F. Hornbein and R.B. Schoene), pp. 645–661. New York: Marcel Dekker.
80. Meeuwsen, T., Hendriksen, I.J.M. and Holewijn, M. (2001). Training induced increases in sea-level anaerobic performance are enhanced by acute intermittent hypobaric hypoxia. European Journal of Applied Physiology, 84, 283–290.
81. Melissa, L., MacDougall, J.D., Tarnopolsky, M.A., Cipriano, N. and Green, H.J. (1997). Skeletal muscle adapta-tions to training under normobaric hypoxic versus normoxic conditions. Medicine and Science in Sports and Exercise, 29, 238–243.
82. Mellerowicz, H., Meller, W., Woweries, J. et al. (1970). Vergleichende Untersuchungen u¨ber Wirkungen von Höhentraining auf die Dauerleistung in Meeresho¨he. Sportarzt und Sportmedizin, 21 (Heft 9–10), 207–240.
83. Miyazaki, S. and Sakai, A. (2000). The effect of ‘living high– training low’ on physical performance in rats. International Journal of Biometeorology, 44, 24–30.
84. Mizuno, M., Juel, C., Bro-Rasmussen, T. et al. (1990). Limb skeletal muscle adaptation in athletes after training at altitude. Journal of Applied Physiology, 68, 496–502.
85. Morris, D.M., Kearney, J.T. and Burke, E.R. (2000). The effects of breathing supplemental oxygen during altitude training on cycle performance. Journal of Science and Medicine in Sport, 3, 165–175.
86. Nielsen, H.B., Bredmose, P.P., Stra˚mstad, M. et al. (2002). Bicarbonate attenuates arterial desaturation during maximal exercise in humans. Journal of Applied Physiology, 93, 724–731.
87. Noakes, T.D. (2000). Physiological models to understand exercise fatigue and the adaptations that predict or enhance athletic performance. Scandinavian Journal of Medicine and Science in Sports, 10, 123–145.
88. Noakes, T.D., Peltonen, J. and Rusko, H. (2001). Evidence that a central governor regulates exercise performance during acute hypoxia and hyperoxia. Journal of Experimental Biology, 204, 3225–3234.
89. Nummela, A.T. and Rusko, H. (2000). Acclimatization to altitude and normoxic training improve 400-m running performance at sea level. Journal of Sports Sciences, 18, 411–419.
90. Paavolainen, L., Nummela, A. and Rusko, H. (1999a). Neuromuscular characteristics and muscle power as determinants of 5-km running performance. Medicine and Science in Sports and Exercise, 31, 124–130.
91. Paavolainen, L., Nummela, A., Rusko, H. and Ha¨kkinen, K. (1999b). Neuromuscular characteristics and fatigue during 10 km running performance. International Journal of Sports Medicine, 20, 516–521.
92. Paavolainen, L., Ha¨kkinen, K., Ha¨ma¨la¨inen, I., Nummela, A. and Rusko, H. (1999c). Explosive-strength training improves 5-km running time by improving running economy and muscle power. Journal of Applied Physiology, 86, 1527–1533.
93. Paavolainen, L., Nummela, A. and Rusko, H. (2000). Muscle power factors and VO2max as determinants of horizontal and uphill running performance. Scandinavian Journal of Medicine and Science in Sports, 10, 286– 291.
94. Peltonen, J., Rantamäki, J., Niittymäki, S. et al. (1995). Effects of oxygen fraction in inspired air on rowing performance. Medicine and Science in Sports and Exercise, 27, 573–579.
95. Peltonen, J., Rusko, H., Rantamäki, J. et al. (1997). Effects of oxygen fraction in inspired air on force production and electromyogram activity during ergometer rowing. European Journal of Applied Physiology, 76, 495– 503.
96. Peltonen, J., Leppa¨vuori, A.P., Kyrö, K.-P., Mäkela¨, P. and Rusko, H. (1999). Arterial hemoglobin oxygen saturation is affected by FiO2 at submaximal running velocities in elite athletes. Scandinavian Journal of Medicine and Science in Sports, 9, 265–271.
97. Peltonen, J., Tikkanen, H. and Rusko, H. (2001). Cardiorespiratory responses to exercise in acute hypoxia, hyperoxia and normoxia. European Journal of Applied Physiology, 85, 82–88.
98. Piehl-Aulin, K. (1999). Normobaric hypoxia – physical performance. Journal of Sports Sciences, 12, 478–479.
99. Piehl-Aulin, K., Svedenhag, J., Wide, L., Berglund, B. and Saltin, B. (1998). Short-term intermittent normobaric hypoxia – hematological, physiological and mental effects. Scandinavian Journal of Medicine and Science in Sports, 8, 132–137.
100. Rahkila, P. and Rusko, H. (1982). Effect of high altitude training on muscle enzyme activities and physical performance characteristics of cross-country skiers. In Exercise and Sport Biology, International Series on Sport Sciences, Vol. 12. (edited by P.V. Komi), pp. 143–151. Champaign, IL: Human Kinetics.
101. Remes, K. (1979). Effect of long-term physical training on total red cell volume. Scandinavian Journal of Clinical and Laboratory Investigations, 39, 311–319.
102. Reynafarje, B. (1962). Myoglobin content and enzymatic activity of muscle and altitude adaptation. Journal of Applied Physiology, 17, 301–305.
103. Reynafarje, C., Lozano, R. and Valdivieso, J. (1959). The polycythemia of high altitudes: iron metabolism and related aspects. Blood, 14, 433–455.
104. Ri-Li, G., Witkowski, S., Zhang, Y. et al. (2002). Determinants of erythropoietin release in response to short-term hypobaric hypoxia. Journal of Applied Physiology, 92, 2361– 2367.
105. Roach, R.C. and Hackett, P.H. (2001). Frontiers of hypoxia research: acute mountain sickness. Journal of Experimental Biology, 204, 3161–3170.
106. Robbins, P.A. (2001). Is ventilatory acclimatization to hypoxia a phenomenon that arises through mechanisms that have an intrinsic role in the regulation of ventilation at sea level. In Hypoxia: From Genes to the Bedside. Proceedings of the 12th International Hypoxia Symposium (edited by R.C. Roach, P.D. Wagner and P.H. Hackett), pp. 339– 348. New York: Kluwer Academic/Plenum Publishers.
107. Roberts, A.D., Clark, S.A., Townsend, N.E. et al. (2003). Changes in performance, maximal oxygen uptake and maximal accumulated oxygen deficit after 5, 10 and 15 days of live high:train low altitude exposure. European Journal of Applied Physiology, 88, 390–395.
108. Rodríguez, F.A., Casas, H., Casas, M. et al. (1999). Intermittent hypobaric hypoxia stimulates erythropoiesis and improves aerobic capacity. Medicine and Science in Sports and Exercise, 31, 264–268.
109. Rodríguez, F.A., Murio, J. and Ventura, J.L. (2003). Effects of intermittent hypobaric hypoxia and altitude training on physiological and performance parameters in swimmers. Medicine and Science in Sports and Exercise, 35, S115.
110. Roskam, H., Landry, F., Samek, L. et al. (1969). Effect of a standardized ergometer training program at three different altitudes. Medicine and Science in Sports and Exercise, 27, 840–847.
111. Rusko, H. (1996). New aspects of altitude training. American Journal of Sports Medicine, 24, S48–S52.
112. Rusko, H. (2003). Handbook of Sports Medicine and Science: Cross Country Skiing. Oxford: Blackwell Science.
113. Rusko, H. and Nummela, A.T. (1996). Measurement of maximal and submaximal anaerobic power. International Journal of Sports Medicine, 17, S89–S130.
114. Rusko, H. and Tikkanen, H. (1996). Normobaric hypoxia and hyperoxia in training and medical rehabilitation. Suomen Lääkärilehti, 31, 3267–3273 (in Finnish).
115. Rusko, H., Penttinen, J.T.T., Koistinen, P.O., Vähäsöyrinki, P.I. and Leppäluoto, J.O. (1995a). A new solution to simulate altitude and stimulate erythropoiesis at sea level in athletes. In The Way to Win. Proceedings of the International Congress on Applied Research in Sports (edited by J. Viitasalo and U. Kujala), pp. 287–289. Helsinki: Liikuntatieteellinen Seura.
116. Rusko, H., Leppävuori, A., Ma¨kela¨, P. and Leppäluoto, J. (1995b). Living high, training low: a new approach to altitude training at sea level in athletes. Medicine and Science in Sports and Exercise, 27, S6.
117. Rusko, H., Kirvesniemi, H., Paavolainen, L., Vähäsöyrinki, P. and Kyro¨, K.-P. (1996). Effect of altitude training on sea level aerobic and anaerobic power of elite athletes. Medicine and Science in Sports and Exercise, 28, S124.
118. Rusko, H., Tikkanen, H., Paavolainen, L. et al. (1999). Effect of living in hypoxia and training in normoxia on sea.
119. Level VO2 max and red cell mass (1985). Medicine and Science in Sports and Exercise, 31, S86.
120. Saltin, B. (1967). Aerobic and anaerobic work capacity at an altitude of 2,250 meters. In Proceedings of a Symposium on the Effects of Altitude on Physical Performance (edited by R.F. Goddard), pp. 97–102. Chicago, IL: The Athletic Institute.
121. Saltin, B., Larsen, H., Terrados, N. et al. (1995a). Aerobic exercise capacity at sea level and at altitude in Kenyan boys, junior and senior runners compared with Scandina-vian runners. Scandinavian Journal of Medicine and Science in Sports, 5, 209–221.
122. Saltin, B., Kim, C.K., Terrados, N. et al. (1995b). Morphology, enzyme activities and buffer capacity in leg muscles of Kenyan and Scandinavian runners. Scandinavian Journal of Medicine and Science in Sports, 5, 222–230.
123. Schmidt, W., Eckardt, K.-U., Strauch, S. and Bauer, C. (1991). Effects of maximal and submaximal exercise under normoxic and hypoxic conditions on serum erythropoietin level. International Journal of Sports Medicine, 12, 457–461.
124. Schmidt, W., Heinicke, K., Rojas, J. et al. (2002). Blood volume and hemoglobin mass in endurance athletes from moderate altitude. Medicine and Science in Sports and Exercise, 34, 1934–1940.
125. Schneider, M., Bernasch, D., Weymann, J., Holle, R. and Bärtsch, P. (2002). Acute mountain sickness: influence of susceptibility, preexposure, and ascent rate. Medicine and Science in Sports and Exercise, 34, 1886–1891.
126. Schoene, R.B. (1982). Control of ventilation in climbers to extreme altitude. Journal of Applied Physiology, 53, 886–890.
127. Schoene, R.B. (2001). Limits of human lung function at high altitude. Journal of Experimental Biology, 204, 3121– 3127.
128. Serebrovskaya, T.V. (2002). Intermittent hypoxia research in the former Soviet Union and the Commonwealth of Independent States: history and review of the concept and selected applications. High Altitude Medicine and Biology, 3, 205–221.
129. Shepley, B., MacDougall, J.D., Cipriano, N. et al. (1992). Physiological effects of tapering in highly trained athletes. Journal of Applied Physiology, 72, 706–711.
130. Squires, R.W. and Buskirk, E.R. (1982). Aerobic capacity during acute exposure to simulated altitude, 914 to 2286 metres. Medicine and Science in Sports and Exercise, 14, 36– 40.
131. Stray-Gundersen, J., Alexander, C., Hochstein, A., de Lemos, D. and Levine, B.D. (1992). Failure of red cell volume to increase to altitude exposure in iron deficient runners. Medicine and Science in Sports and Exercise, 24, S90.
132. Stray-Gundersen, J., Hochstein, A. and Levine, B.D. (1993). Effect of 4 weeks altitude exposure and training on red cell mass in trained runners. Medicine and Science in Sports and Exercise, 25, S171.
133. Stray-Gundersen, J., Mordecai, N. and Levine, B.D. (1995). O2 transport response to altitude training in runners. Medicine and Science in Sports and Exercise, 27, 202.
134. Stray-Gundersen, J., Levine, B.D. and Bertocci, L.A. (1999). Effect of altitude training on runners’ skeletal muscle. Medicine and Science in Sports and Exercise, 31, S182.
135. Stray-Gundersen, J., Karlsen, T., Resaland, G.K. et al. (2000). No difference in 3-day EPO response to 8, 12 or 16 hours/day of intermittent hypoxia. Medicine and Science in Sports and Exercise, 32, S251.
136. Stray-Gundersen, J., Chapman, R. and Levine, B.D. (2001). ‘Living high–training low’ altitude training improves sea level performance in male and female elite runners. Journal of Applied Physiology, 91, 1113–1120.
137. Svedenhag, J., Saltin, B., Johansson, C. and Kaijser, L. (1991). Aerobic and anaerobic exercise capacities of elite middle-distance runners after two weeks of training at moderate altitude. Scandinavian Journal of Medicine and Science in Sports, 1, 205–214.
138. Svedenhag, J., Piehl-Aulin, K., Skog, C. and Saltin, B. (1997). Increased left ventricular muscle mass after long-term altitude training in athletes. Acta Physiologica Scandinavica, 161, 63–70.
139. Terrados, N., Mizuno, M. and Andersen, H. (1985). Reduction in maximal oxygen uptake at low altitudes: role of training status and lung function. Clinical Physiology, 5 (suppl. 3), 75–79.
140. Terrados, N., Melichna, J., Sylven, C., Jansson, E. and Kaijser, L. (1988). Effects of training at simulated altitude on performance and muscle metabolic capacity in competitive road cyclists. European Journal of Applied Physiology, 57, 203–209.
141. Terrados, N., Jansson, E., Sylven, C. and Kaijser, L. (1990). Is hypoxia a stimulus for synthesis of oxidative enzymes and myoglobin? Journal of Applied Physiology, 68, 2369– 2372.
142. Tkatchouk, E.N., Gritsenko, P.V., Tsvetkova, A.M., Zhou, Z.-N. and Wu, X.F. (1999). Oxygen transport in persons adapted to hypoxia during acute hypobaric hypoxia. In Hypoxia: Into the Next Millennium. Proceedings of the 11th International Hypoxia Symposium (edited by R.C. Roach, P.D. Wagner and P.H. Hackett), pp. 434–435. New York: Kluwer Academic/Plenum Publishers.
143. Townsend, N.E., Gore, C.J., Hahn, A.G. et al. (2002). Living high–training low increases hypoxic ventilatory response of well-trained endurance athletes. Journal of Applied Physiology, 93, 1498–1505.
144. Vasankari, T., Rusko, H., Kujala, U.M. and Huhtaniemi, I.T. (1993). The effect of ski training at altitude and racing on pituitary, adrenal and testicular function in men. European Journal of Applied Physiology, 66, 221–225.
145. Vasankari, T.J., Kujala, U.M., Rusko, H., Sarna, S. and Ahotupa, M. (1997). The effect of endurance exercise at moderate altitude on serum lipid peroxidation and antioxidative functions in humans. European Journal of Applied Physiology, 75, 396–399.
146. Ventura, N., Hoppeler, H., Seiler, R. et al. (2003). The response of trained athletes to six weeks of endurance training in hypoxia or normoxia. International Journal of Sports Medicine, 24, 166–172.
147. Wagner, P.D. (2000). New ideas on limitations to VO2max.
148. Exercise and Sport Sciences Reviews, 28, 10–14. Ward, M.P., Milledge, J.S. and West, J.B. (1999). High.
149. Altitude Medicine and Physiology (1989). New York: Oxford.
150. University Press. Wehrlin, J., Zuest, P., German, C., Hallen, J. and Bernard, M. (2003). 24 days live high–train low increases red cell volume, running performance and VO2max in Swiss national team orienteers. In Book of Abstracts, 8th Annual Congress of the European College of Sport Science (edited by E. Müller, H. Schwameder, G. Zallinger and V. Fastenbauer), pp. 375–376. Salzburg: Institute of Sport Science, University of Salzburg.
151. Weil, J.V., Jamieson, G., Brown, D.W. and Grover, R.F. (1968). The red cell mass–arterial oxygen relationship in normal man. Journal of Clinical Investigation, 47, 1627– 1639.
152. Wilber, R.L. (2001). Current trends in altitude training. Sports Medicine, 31, 249–265.
153. Wilber, R.L. (2003). Effect of FiO2 on physiological responses and cycling performance at moderate altitude. Medicine and Science in Sports and Exercise, 35, 234–236.
154. Wilber, R.L., Drake, S.D., Hesson, J.L. et al. (2000). Effect of altitude training on serum creatine kinase activity and serum cortisol concentration in triathletes. European Journal of Applied Physiology, 81, 140–147.
155. Wolfel, E.E., Groves, B.M., Brooks, G.A. et al. (1991). Oxygen transport during steady-state submaximal exercise in chronic hypoxia. Journal of Applied Physiology, 70, 1129– 1136.
Cita Original
Heikki Rusko, Heikki Tikkanen and Juha Peltonen (2004) Altitude and endurance training. Journal of Sports Sciences, 22:10, 928-945, DOI: 10.1080/02640410400005933
Cita en PubliCE
Heikki Rusko, Heikki Tikkanen Juha Peltonen (2016). Altura y Entrenamiento de Resistencia. .https://g-se.com/altura-y-entrenamiento-de-resistencia-2091-sa-t57cfb2727a029