Actividad Física, Gasto de Energía y Aptitud Física: Una Perspectiva Evolutiva
R. W Gotshall1, L Cordain1, Boyd Eaton2, Boyd Eaton III31 Department of Exercise and Sport Science, Colorado State University. Fort Collins CO, Estados Unidos.
2 Department of Radiology, Emory University School of Medicine, Atlanta CA, Estados Unidos.
3 Department of Education, Marshall University. Huntington. WV, Estados Unidos.
Resumen
El modelo de los patrones de actividad física de los seres humanos no fue establecido en los gimnasios, campos de deporte ni en los laboratorios de fisiología de ejercicio, si no que a través de la selección natural que actúa a lo largo de los eones de experiencia evolutiva. Este trabajo analiza cómo la evolución ha determinado el potencial para el rendimiento del humano contemporáneo y propone la experiencia de los cazadores recolectores estudiados recientemente como el mejor indicador disponible (aunque reconocidamente imperfecto) de los patrones de actividad física para los cuales nuestra biología, genéticamente determinada, fue seleccionada originalmente. Desde la aparición del género Homo, hace aprox. 2 millones de años (MYA), hasta la revolución agrícola de aproximadamente 10000 años atrás, nuestros antepasados eran cazadores recolectores, por lo que las presiones adaptativas inherentes a ese nicho medioambiental ejercieron influencia definitiva en la composición genética humana. La porción de nuestro genoma que determina la anatomía y la fisiología básica, ha permanecido relativamente inalterada durante los últimos 40000 años. Así, la relación mutua compleja entre la ingesta de energía, gasto de energía y los requisitos de actividad física específicos para los seres humanos actuales, siguen siendo muy similares a los seleccionados originalmente para los hombres y mujeres de la Edad de Piedra que vivieron de la recolección y la caza. Las investigaciones que analicen la actividad física óptima para la salud y el rendimiento humano pueden guiarse a través del conocimiento de la evolución de los patrones de actividad física en nuestra especie.
Palabras Clave: ejercicio, evolución, aptitud física, antropología, salud, selección natural
INTRODUCCION
El modelo de los patrones de actividad física de los seres humanos no se estableció en los gimnasios, campos de deportes o laboratorios de fisiología de ejercicio, si no que fue establecido por la selección natural, que actúa a lo largo de los eones de experiencia evolutiva. Este trabajo analiza cómo la evolución ha determinado el potencial del rendimiento humano contemporáneo y aborda la experiencia de los cazadores recolectores recientemente estudiado como el mejor indicador disponible (aunque reconocidamente imperfecto) de los patrones de actividad física para los cuales nuestra biología genéticamente determinada fue originalmente seleccionada. La información revisada en este informe es el resultado de una extensa búsqueda bibliográfica utilizando Medline, Sport Discus y Colorado Alliance of Research Libraries y también las bibliografías de artículos originales.
Virtualmente toda la literatura biomédica que considera la frecuencia, intensidad, duración y tipo de ejercicio óptimos (por ejemplo 36) abordan los mecanismos inmediatos que relacionan el ejercicio con la salud y la aptitud física; sin embargo, la perspectiva evolutiva se focaliza en las bases principales para estas relaciones (17). Aunque hay variación individual sustancial en las capacidades de realizar ejercicio debido a diferentes factores de desarrollo y comportamiento, y debido a la diversidad genética individual, el rango funcional humano final está genéticamente determinado (17). Al igual que los otros organismos, nuestros antepasados desarrollaron capacidades, limitaciones y requisitos físicos específicos de la especie a medida que competían en sus nichos medioambientales y, durante dos millones de años hasta la aparición de la agricultura hace sólo 10000 años (Tabla 1), los antepasados humanos eran cazadores recolectores 1 El estrés profesional y medioambiental de este estilo de vida formaron el conjunto de genes ( pool génico) del humano contemporáneo.
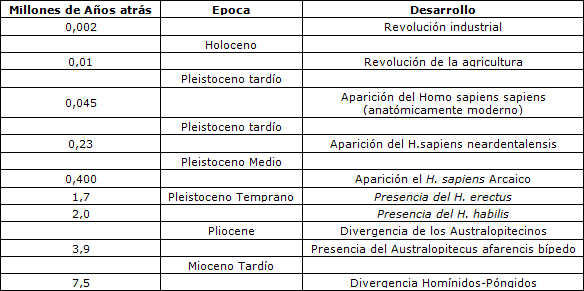
Tabla 1. Principales eventos de evolución humana.
Si bien anatómicamente los humanos modernos aparecieron primero hace quizás 100000 años, el comportamiento humano moderno fue reconocido por primera vez en el registro fósil aproximadamente 50000 años atrás. Las comparaciones del ADN mitocondrial de los diferentes grupos étnicos humanos (58,62) indican que la constitución genética de hombres y mujeres contemporáneos ha cambiado relativamente poco durante estos 50 milenios a pesar de los enormes cambios sociales asociados con la agricultura y la industrialización. Así la relación entre la ingesta de energía, gasto de energía y la actividad motora específica, todavía sigue siendo la que originalmente fue seleccionada para la vida en la edad de piedra, en un ambiente donde se vivía forrajeando. Sin embargo, los elementos que facilitan las tareas domésticas y laborales, el transporte motorizado y las actividades recreativas cada vez más sedentarias, han reducido la cantidad de ejercicio físico obligatorio, muy por debajo del nivel obtenido cuando se seleccionó el genoma humano existente. En las sociedades industrializadas, la actividad física para la mayoría de las personas se ha vuelto una actividad extraordinaria, principalmente separada de otras tareas diarias, y realizada específicamente para mejorar las variables de aptitud física como la resistencia, fuerza y/o flexibilidad. De manera contraria, el ejercicio físico diario era un aspecto obligatorio integral, de la existencia de nuestros antepasados: la capacidad de cazar, recolectar, cargar, excavar y escapar de los depredadores dependía de la actividad muscular y de la aptitud física individual.
Cambios Evolutivos en la Anatomía y Fisiología de los Homínidos que Influyeron en la Capacidad de Realizar Ejercicios
Aunque el origen de los mamíferos se extendió finalmente al principio de la Era Mesozoica, nuestros ancestros prehumanos (los homínidos) y los de los grandes monos de la actualidad (póngidos) divergieron relativamente recientemente, hace quizás 5-7 millones de años (MYA). Las influencias evolutivas que nos hacen singularmente humanos han operado principalmente durante el período subsecuente a esta división entre homínidos y póngidos. Los primeros homínidos conocidos, con una locomoción vertical indiscutible fueron los Australopitecinos, quienes aparecen en el registro fósil africano oriental entre 4,2 y 3,9 MYA (29). Durante el mismo período de tiempo, los cambios climáticos redujeron la magnitud de los bosques tropicales y llevaron a la formación de un ambiente más, seco más abierto con forma de mosaico entre selva y sabana (21). En este contexto tener una postura erguida y una locomoción bípeda les confirió algunas ventajas: ayudó a la localización visual de comida, agua y depredadores (50); liberó las manos para cargar, excavar y utilizar armas (35); y minimizó el área de superficie corporal expuesta al sol, lo que compensó parcialmente la carga térmica que resultaba de la actividad física diaria (14, 59, 61).
Cuando los humanos y otros primates corren. Sus costos de energía son aproximadamente dos veces superiores al de otros mamíferos (55, 56), pero la locomoción humana es tan eficaz como la locomoción cuadrúpeda (45, 57). Las piernas humanas tienen una estructura similar a una palanca de apoyo que les permite transferir la energía potencial y cinética de una manera más eficiente que las piernas de los mamíferos cuadrúpedos (12,13). Estas consideraciones ayudan a explicar por qué nuestros antepasados Australopitecinos pudieron volverse bípedos; Tuvieron un impacto profundo en las características físicas y en el potencial de rendimiento de sus descendientes, incluyéndonos a nosotros mismos.
El primer Australopitecino, el A. afarensis, retuvo muchas características estructurales de los miembros superiores similares a las de los monos (lo que les permitía un continuo acceso a los árboles para alimentarse, descansar y estar seguros [53]), pero el análisis de las huellas fósiles y de las estructuras óseas de la pelvis/piernas indica que caminaban y corrían con una eficacia mecánica bastante similar a la de los humanos contemporáneos (35). Ellos eran mucho más pequeños que nosotros: los varones adultos medían aproximadamente 1,5 m, pesaban aproximadamente 45 kg y sus capacidades craneales eran sólo 400-500 mL (1) (Figura l). Al igual que los póngidos vivientes, tenían más pelo en el cuerpo que el que tienen los humanos actuales, aunque la reducción del pelo corporal, para mejorar la dispersión de calor, se produjo probablemente con el tiempo. Esta adaptación actuó en forma conjunta con la locomoción bípeda para permitir una mayor actividad física durante el día (60). La pérdida de pelo sólo puede reducir la carga térmica eficazmente si se presenta junto con un buen sistema de glándulas sudoríparas cutáneas (o ecrinas), capaces de realizar un enfriamiento por evaporación (40). Las glándulas sudoríparas del humano actual pueden producir más sudor por área de superficie que las de cualquier otra especie de mamífero (11, 40), una tasa máxima de dispersión de calor en exceso de 500 Watts/metro2 (la tasa máxima para los primates modernos con pelo es 100 Watts/metro2) (60). La disminución en el pelo y un complejo sistema de glándulas sudoríparas que facilite la dispersión de calor habrían tenido valor adaptativo principalmente si los Australopitecinos eran más activos físicamente durante el día que sus antepasados primates. Esto implica que la caminata, la carrera y otras actividades generadoras de calor se volvieron aspectos importantes de su existencia y que, igualmente, eran incluso posibles durante períodos extensos bajo el intenso sol africano.
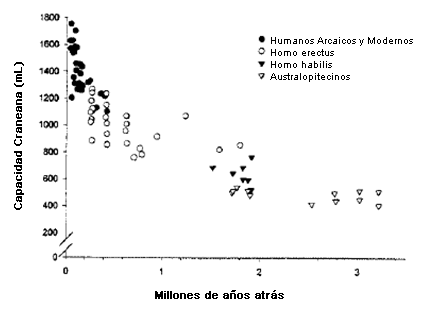
Figura 1. Aumento de la capacidad craneal absoluta en los homínidos a lo largo del tiempo [Adaptado de (1)].
Los primeros Australopitecinos eran capaces de caminar erguidos y probablemente tenían un primer sistema termorregulatorio eficaz. Sus proporciones corporales eran similares a las de los monos, con brazos relativamente más largos y piernas relativamente más cortas que las de sus descendientes, los humanos tempranos (53), quienes poseen huesos del antebrazo (radio y cúbito) más cortos y fémures más largos (26). Por lo tanto las adaptaciones evolutivas subsecuentes al A.afarensis favorecieron una caminata energéticamente más eficiente: la reducción relativa de la masa supraacetabular a través de la disminución en la longitud de los brazos, mejoró el equilibrio mientras que un fémur relativamente más largo facilitó la transferencia de energía potencial y cinética, aumentando así la recuperación de energía mecánica durante la locomoción (11). Por otro lado, la disminución en la longitud del antebrazo tuvo el efecto indeseable de reducir la fuerza máxima de los miembros superiores porque disminuyó la acción la palanca biomecánica del brazo (4, 19, 26).
Los chimpancés se alimentan principalmente de plantas (94%), sobre todo de frutas; el resto de su dieta consiste en insectos y vertebrados pequeños (37). Los paleoantropólogos asumen que los hominoides2 pre-Australopitecinos y los primeros Australopitecinos tenían un modelo de subsistencia similar. Sin embargo, hacia el fin del Plioceno, cuando el bosque tropical se transformó en bosque más abierto y sabana, la disponibilidad de frutas se volvió más estacional. Esos Australopitecinos que fueron los antepasados humanos, respondieron incluyendo más material animal en sus dietas (37); otros homínidos (por ejemplo A. robustus) reaccionaron al mismo desafío medioambiental utilizando alimentos vegetales de menor calidad, desarrollando mandíbulas y molares más macizos para procesar la vegetación más dura. Estas especies de Australopitecinos con una masticación más robusta se extinguieron, pero las líneas proto-humanas, que consumían ahora más alimentos animales, gradualmente evolucionaron hacia la primera especie de nuestro género, el Homo habilis qué era similar en tamaño corporal al A. afarensis pero con un cerebro significativamente mayor (Figura l). El Homo habilis probablemente fue el primer homínido en hacer herramientas de piedra y por lo tanto, se cree que dependía más de la caza y de la recolección. El aumento resultante en la disponibilidad de alimentos de origen animal fue un factor importante en la evolución de un cerebro más grande y de una mayor complejidad comportamental (1).
La evolución de un cerebro más grande, metabólicamente más activo requirió un aumento en la calidad de la dieta (es decir en la densidad calórica) (1, 33). Los humanos destinan 20-25% de su requerimiento metabólico en reposo (RMR) a la alimentación del cerebro, mientras que los primates no humanos sólo necesitan aproximadamente 8% y otros mamíferos sólo 3-4% (32). Sin embargo, el aumento en el metabolismo del cerebro en los primates encefalizados, y especialmente en los humanos, evolucionó sin aumentar el RMR (1), un desarrollo que requirió una disminución equilibrada en el metabolismo de otros órganos del cuerpo. La mayor calidad de la dieta permitió la reducción en el tamaño y en la actividad metabólica del intestino, lo que compensó bastante el aumento en el metabolismo del cerebro (32, 33, 37).
La encefalización se presentó junto con un aumento en la complejidad conductual (47), un mayor rango diario (32), mayores gastos de energía total diarios (TEE) (Tabla2) y una aumento en la estatura: en 1,7 MYA el H. habilis habría evolucionado hacia el Homo erectus, una especie tan alta como los humanos contemporáneos (10,46). El espesor cortical y el diámetro global de sus restos óseos (Figura 2) indican que el H. erectus era muy musculoso (47); en promedio deben haber sido ciertamente más fuertes que la mayoría de los humanos contemporáneos. Sus instrumentos de piedra eran mucho menos sofisticados que los de los humanos posteriores. Ellos no utilizaron herramientas con mango, o palancas compuestas para multiplicar la fuerza, por lo que la carga muscular para estos antepasados humanos tempranos debe haber sido alta. Además de las extremidades musculares, el H. erectus contaba muy probablemente con sistemas cardiovascular, metabólico y termo regulatorio capaces de mantener un ejercicio aeróbico de alto nivel (1,46), adaptaciones fisiológicas necesarias para los homínidos diurnos que viajaban distancias considerables porque cazaban, eran carroñeros, recolectaban y trasladaban cargas en los calurosos climas ecuatoriales.
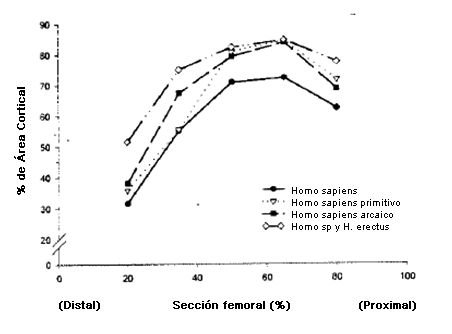
Figura 2. Diferencias en el área cortical por ciento ([área cortical/área de periostio] x 100) a lo largo de la longitud del cuerpo femoral en individuos del género Homo primitivos y actuales (de 47).
El Homo erectus comenzó a extenderse desde África por lo menos hace un millón de años, colonizando las regiones más calurosas del sur de Asia. Las formas intermedias entre el Homo erectus y el Homo sapiens aparecieron quizás hace 500000 a 400000 años. En Europa éstos evolucionaron en los Neardentales. En África originaron al anatómicamente moderno Homo sapiens, que se encuentra presente en el registro fósil un poco antes de 100000 BP (Antes del Presente). Al principio esta modernidad anatómica no se asoció con adelantos culturales arqueológicamente reconocibles: Los H. sapiens vivieron como sus predecesores. Sin embargo, hace 50000 y 40000 años, su creatividad y su innovación tecnológica aumentaron marcadamente (54), de modo que mejoraron la eficacia para cazar, recolectar, procesar los alimentos y muchas otras actividades necesarias. Durante este período la robustez de los esqueletos disminuyó un poco, lo que sugiere que se requirió menos fuerza muscular para las tareas diarias (47). Todavía, los Cromañones y otros humanos anatómicamente modernos de esta era siguieron siendo cazadores recolectores; la actividad física vigorosa siguió siendo un rasgo esencial de su existencia. Un estilo de vida basado en la recolección y la caza siguió siendo el eslabón fundamental entre quienes habitaban en la era de piedra hace 25000 años y aquellos que se dedicaban a forrajear estudiados en este siglo. Las circunstancias en el Paleolítico tardío (35000 a 15000 BP) difirieron de manera importante de las experimentadas por los actuales cazadores recolectores. Por ejemplo las últimas áreas ahora habitadas con disponibilidad marginal de recursos, probablemente provocó su pequeña estatura (que contrastaba con la de los humanos del Paleolítico tardío). No obstante los forrajeadores como las tribus Ache, !Kung, Agta, Hadza e Inuit nos aportan una visión sobre las formas de vida de nuestros antepasados previos a la agricultura, incluyendo visiones sobre los modelos de actividad física para los cuales nuestra especie está constituida genéticamente.
Gasto de Energía, Actividad Física y Aptitud Física
La ingesta de energía diaria se destina al ejercicio físico, crecimiento, reproducción, RMR y a contrarrestar los efectos térmicos de la alimentación (masticación, mayor peristaltismo, digestión, etc.). Cualquier exceso se almacena en forma de grasa. En la actualidad, para los humanos promedio en las naciones adineradas el RMR comprende aproximadamente 70% del TEE y el efecto térmico de la comida 5-10%; la mayoría del TEE restante se atribuye a la actividad física (9). El RMR está estrechamente relacionado con el tamaño corporal en las especies de mamíferos, siendo frecuentemente estimado como 70 veces la masa corporal (en kg) a la 3/4 ta potencia (27).
Patrones de Actividad Física
Antes de este siglo, los patrones de actividad de humanos/prehumanos y los rangos diarios estaban principalmente regulados por la provisión de alimentos. Como la mayoría de los otros primates los Australopitecinos vivieron en hábitats dominados por bosques y selvas. En estas áreas, los alimentos estaban fácilmente disponibles y un rango limitado era suficiente para la subsistencia (Tabla 2). Sin embargo, entre 2,5 y 1,5 MYA, los bosques de África oriental y del sur disminuyeron notablemente debido a un prolongado período de sequía (5, 15). Este cambio medioambiental de bosque a sabana abierta alteró la abundancia y distribución de los recursos alimenticios (22) y requirió que los primeros miembros del género Homo extendieran su rango diario (20). El H. erectus podría haber transportado herramientas, armas y haber cazado en un rango de 15 km (23, 28), al igual que los forrajeadores actuales que tienen rangos diarios sustanciales (Tabla 2).
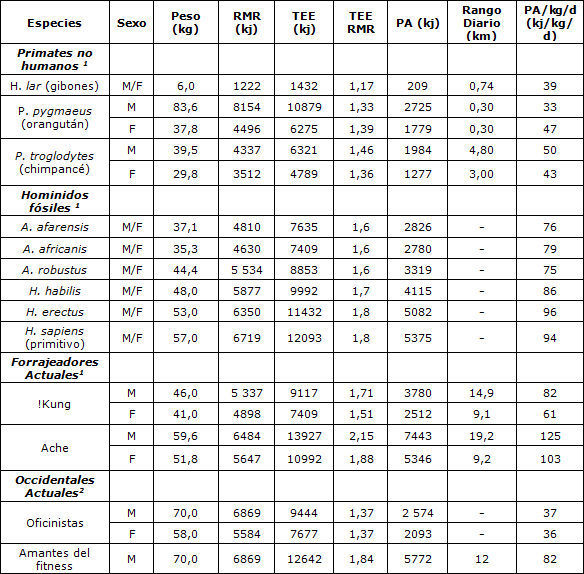
Tabla 2. Gasto de energía de los primates. 1 Obtenido de (32). 2 Obtenidos de National Research Council Recommended Daily Allowances Tablas 3-1 y 3-3 (39). Los amantes del fitness realizan un ejercicio equivalente a correr 7,5 mph, 60min/día(25). RMR = Tasa metabólica en reposo: TEE= Gasto de energía total: PA= Actividad física (sin tener en cuenta el efecto térmico de la comida). 1 kj =0,238 kcal.
El patrón de actividad semanal de cazadores recolectores de este siglo es similar a lo que se denominó Ritmo Paleolítico: los varones normalmente cazan de uno a cuatro días no consecutivos por semana con los días intermedios de descanso y las mujeres rutinariamente realizan recolección cada dos o tres días (48). Otras actividades comunes con demandas físicas frecuentes, incluyen la fabricación de herramientas, cuidado de los hijos (un niño promedio debe ser cargado 1500 km durante sus primeros dos años de vida [30]), matar presas y preparación de otras comidas, preparación de la vestimenta, cargar leña para encender fuego, cargar agua y desplazarse a nuevos sitios de acampe. Los bailes (que pueden extenderse por horas) son una actividad recreativa importante de muchas culturas, y generalmente se realizan varias noches por semana. En conjunto, el Ritmo Paleolítico involucra días de ejercicio físico bastante intenso alternados con días de descanso y actividad liviana (48).
Gasto de Energía Total
Dado que sus rangos diarios eran casi ciertamente mayores, el H. habilis y H. erectus tenían un TEE probablemente mayor que el de los Australopitecinos. Para explicar las diferencias en el tamaño corporal, es útil discutir la relación TEE/RMR en lugar del TEE absoluto. Esta relación ha sido estimada en 1,6 para los Australopitecinos. 1,7 para H. habilis y 1,8 para el H. erectus y el H. sapiens primitivo (32) (Tabla 2). La relación TEE/RMR determinada para los forrajeadores recientemente estudiados se acerca a 2,0 (para los agricultores rurales contemporáneos puede superar el valor de 2,0), pero en contraste notable, la relación TEE/RMR para los humanos modernos, sedentarios en las sociedades ricas está por debajo de 1,4 (Tabla 2).
La Figura. 3 presenta el gasto de energía estimado como una función de masa corporal y sugiere que RMR y TEE, expresados por kg de peso corporal, se mantuvieron relativamente estables en los antepasados humanos a lo largo de un período de 3,5 millones de años hasta que los H. sapiens contemporáneos se volvieran ricos y sedentarios. Los occidentales actuales típicos tienen valores de TEE/kg/d prácticamente equivalentes al valor de RMR/kg/d de los cazadores recolectores actuales y del estimado para nuestros antepasados preagricultores. La reducción de RMR/kg/d en los humanos modernos probablemente refleja una alteración en la composición corporal (más grasa y menos músculo), un resultado de la vida sedentaria (43). El TEE/kg/d de los humanos contemporáneos típicos es aproximadamente 65% del de los Paleolíticos tardíos de la edad de piedra (asumiendo que su TEE/kg/d era similar al de los forrajeadores actuales; 134 kj/kg/d para los humanos contemporáneos comparados con el promedio de 206 kj/kg/d para los !Kung y los Ache). Cuando se resta RMR del TEE se obtiene una medida bruta del gasto de energía debido a la actividad física (PA) (aunque no se consideran los efectos térmicos de la comida). El gasto de energía por unidad de masa corporal de PA para los occidentales contemporáneos es aproximadamente 38% del de nuestros antepasados humanos, una diferencia aun más extraordinaria que la del TEE. Para que los americanos típicos se aproximen al TEE/kg/d de cazadores recolectores recientemente-estudiados deberían sumar aprox. 72 kj/kg/d (aprox.17 kcal/kg/d), el equivalente de una caminata de 19 km (12 millas) para un hombre de 70 kg, al nivel de actividad física actual por día.
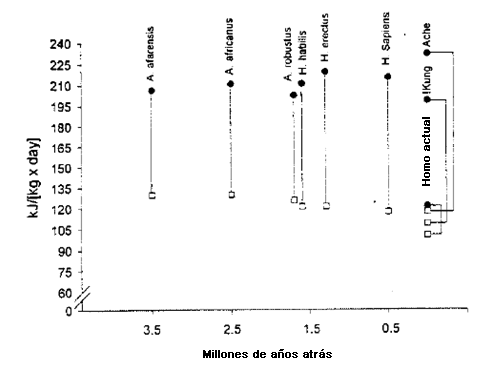
Figura 3. Estimaciones del gasto de energía diario de los Homínidos a lo largo de los últimos 3,5 millones de años (adaptado de [32] y tabla 2). Los cuadrados abiertos son las tasas metabólicas en descanso; los círculos sólidos son los gastos de energía totales. Los valores para los indios mexicanos están ponderados por el ritual de carrera de los corredores Tarahumara.
Niveles de Aptitud Física Aeróbica
No debería sorprendernos que la limitada actividad física típica de los humanos ricos modernos genere aptitud aeróbica mediocre, ni que los niveles aptitud aeróbica de los forrajeadores recientemente estudiados sea superior a la de norteamericanos. Los cazadores recolectores y otras poblaciones tradicionales tienen niveles de aptitud aeróbica que van de buena a excelente cuando son graficadas contra los modelos de aptitud derivados de los estudios de Americanos (49, 51) (Figura 4). Los efectos de la modernización también son predecibles. Entre 1970 y 1990 una comunidad de Inuit canadienses se transformó desde un estilo de vida parcialmente tradicional (cazadores/pescadores) hacia un modo de vida principalmente occidental que incluía el uso extenso de móviles de esquí, motores externos y equipos que facilitaban las labores domésticas (44). Durante un período de 20 años los Inuit se volvieron más gordos como grupo y perdieron fuerza muscular y aptitud física aeróbica. En 1970, antes de la transformación cultural, hombres y mujeres Inuit tenían mayor aptitud física que los hombres de las naciones industrializadas de edad similar, pero para 1990, la superioridad física de los Inuit se había vuelto mucho menos pronunciada (Figura 5).
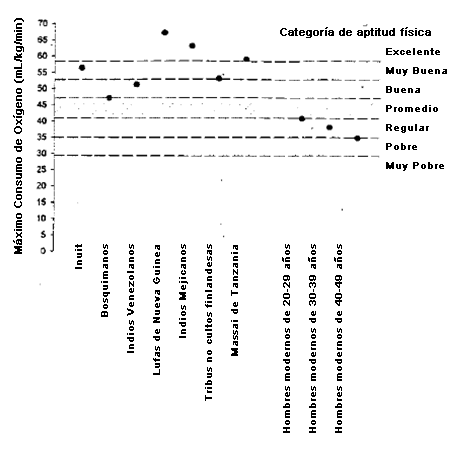
Figura 4. Comparaciones de capacidades aeróbicas máximas (consumo de oxígeno máximo) entre las sociedades no culturizadas y culturizadas (industrializadas) (adaptado de [18]). Las categorías de aptitud física están basadas en (51) las sociedades modernas y transformadas.
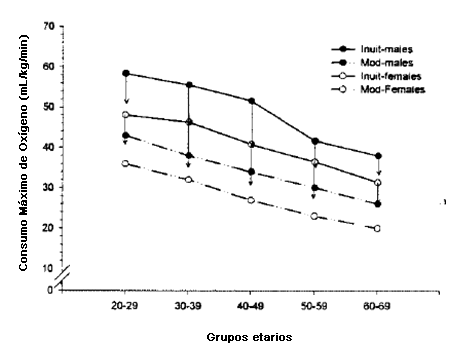
Figura 5. Comparaciones entre las capacidades aeróbicas máximas (consumo de oxígeno máximo) entre los Inuit y las poblaciones no culturizadas por grupos etarios (adaptado de [44]) para los Inuit, y de [51] para las poblaciones culturizadas). Mod = Población moderna culturizada. Las flechas hacia abajo representan el cambio en la aptitud física de los Inuit a lo largo de 20 años de culturización producida de 1970 a 1990 (44).
Recomendaciones Actuales para la Aptitud Física y Salud
Existe mucho interés acerca de la cantidad de actividad física que debe ser recomendada por los profesionales de la salud. La actividad/entrenamiento físico mejora el rendimiento deportivo y la salud general, beneficios que no son mutuamente excluyentes. Las características del rendimiento físico han sido definidas claramente: las recomendaciones del Colegio Americano de Medicina del Deporte (ACSM) de 1995 (2) recomiendan realizar actividad física 3-5 días/semana, al 50-85% de intensidad máxima, continuamente durante 20 a 60 minutos. Sin embargo, para mejorar la salud, el ACSM sugiere que los individuos acumulen 30 minutos de actividad física durante la mayoría de los días de la semana (2). La actividad necesariamente debe ser continua, y puede ser de intensidad moderada (equivalente a caminar 4,83 kilómetros por hora, o 3 millas por hora). Por otro lado, varios informes recientes han puesto énfasis en la naturaleza de la dosis-respuesta de la actividad física y los resultados relacionados con la salud (6, 8, 24, 42).
Al comparar los requisitos de actividad física mínima para mejorar la salud, tal como los propone el ACSM (2) con las estimaciones de ejercicio físico ancestrales, se observa una diferencia llamativa. Un hombre de 70 kg que camina 4,83 km por hora durante 30 min (1,5 mi) gasta aprox 628 kj adicionales (aprox. 150 kcal). Agregando estos 628 kj a los aprox. 2574 kj (615kcal) que un típico trabajador de oficina varón gasta en las actividades diarias (Tabla 2), se obtiene un total de 3202 kj/d (45,7 kj/kg/d, o 11 kcal/kg/d), un valor que cumple con las recomendaciones de ACSM. Sin embargo, este gasto está muy por debajo de los 82 kj/kg/d (19,6 kcal/kg/d) de los varones !Kung o de los varones Ache, que gastan 125 kj/kg/d (24,7 kcal/kg/d). Las recomendaciones de ACSM son aproximadamente 44% del nivel de energía gastados por PA, observado entre cazadores recolectores3, ciertamente muy por debajo de los niveles de nuestros antepasados pre agriculturales y, muy probablemente, por debajo del nivel de ejercicio físico para el cual, nuestra fisiología y bioquímica han sido programadas genéticamente a través de la evolución.
El gasto de energía total de la actividad física puede ser un componente importante para los beneficios sobre la salud (7,31). Los gastos de energía relacionados a la actividad semanal de 4186 a 12558 kj (1000 a 3000 kcal) han sido los propuestos como requisito para obtener beneficios significativos para la salud (38,41,52). Para un hombre de 70 kg esto implica 63 a 179 kJ/kg/semana (15 a 45 kcal/kg/semana), muy por debajo del gasto de los forrajeadores modernos, ya que los varones !Kung gastan 573 kj/kg/sem (137 kcal/kg/sem) y los Ache 724 kj/kg/sem (173 kcal /kg/sem), y también muy por debajo de las estimaciones para los antepasados humanos que vivieron en épocas previas a la agricultura.
Si el nivel de actividad física sugerido por el ACSM para mejorar la salud puede ser considerado un mínimo, ¿qué nivel es necesario para aumentar los beneficios para la salud?. La respuesta estaría en la experiencia de nuestros antepasados y, en ese caso, podrían ser gastos de energía de PA en el rango de 377 kj/kg/sem (90 kcal/kg/semana); el equivalente a caminar 406 kilómetros (252 millas) por mes además de las actividades físicas actuales. Si bien esto parece extremo, el rango diario de los varones !Kung es aproximadamente 15 kilómetros (9 millas) por día, una figura cercana a la de William Wordsworth cuya experiencia puede servir como un ejemplo de niveles de actividad justo antes de que comience la era Industrial y quién se estima que caminó 289800 kilómetros (180000 millas) durante su vida (aproximadamente 15 kilómetros/día durante 60 de sus 70 años)! (16). En la perspectiva evolutiva es la existencia sedentaria la que caracteriza la vida en las naciones ricas contemporáneas lo que representa el extremo, no el estilo de vida que prevaleció en la mayoría de los humanos desde el origen de nuestro género, hasta bien entrada la industrialización 200 años después.
Conclusiones
Con la excepción de Homo sapiens, los mamíferos tienen que trabajar para comer: la procuración del alimento depende directamente del gasto de energía. Sin embargo, los avances tecnológicos y la organización social han roto esta relación básica en los humanos contemporáneos. En las naciones ricas de hoy no existe más un vínculo obligatorio entre la comida que comemos y la energía que gastamos. La energía de los alimentos se ha vuelto mas económica y accesible, mientras que la mecanización ha reducido el ejercicio físico asociado con el trabajo y las actividades recreativas se han vuelto más sedentarias. El resultado es un sobrante relativo de energía, aumento en el peso corporal, sarcopenia y distorsión en la composición corporal con un exceso de tejido adiposo en relación al hueso y músculo (18, 34). La evolución ha dotado a los humanos de una adaptabilidad excepcional, pero nuestra capacidad en este sentido tiene límites y las circunstancias actuales, noveles en la experiencia zoológica evolutiva y comparativa, parecen haber excedido nuestro umbral. La biología humana se ha vuelto tan desordenada que los factores de riesgos fisiológicos y bioquímicos que afectan al sistema cardiovascular, al esqueleto y a nuestro metabolismo de carbohidratos, son en la actualidad increíblemente comunes. En cada caso, el alejamiento de los patrones de ejercicio que prevalecieron durante la evolución tiene un papel fundamental (18).
Per-Olof Ástrand declaró que, "A través de los siglos los humanos han seleccionado una ingesta de comida suficiente para aportar una ingesta de energía de 12000 kj (aproximadamente 3000 kcal) o más" (3). Esta discusión se une a datos paleontológicos que indican que un TEE de 209 kj/kg/d (50 kcal/kg/d) o 14,651 kj/d (3500 kcal) para un humano de70 kg era típico de los antepasados humanos a partir de la aparición de A. afarensis hasta finales del siglo 19. Las circunstancias de vida en las naciones ricas, industrializadas de hoy, son escasamente conducentes a agregar ejercicio físico con otros 4186-6279 kj (1000-1500kcal) al agitado ritmo de cada día. Sin embargo, las actividades de nuestros antepasados, aunque eran exigentes, carecían de la eficacia del ejercicio físico realizado según los principios de la fisiología del ejercicio moderna. Puede ser posible lograr efectos fisiológicos similares con menos consumo de tiempo: se necesitan investigaciones diseñadas para explorar esta posibilidad. Imitar los beneficios fisiológicos y metabólicos esenciales del ejercicio físico, tal como se experimentó a lo largo de la evolución de la humanidad, es una meta lógica y atractiva cuyo logro podría permitir mejorar la salud, prevenir enfermedades y alcanzar el bienestar general. Los métodos más eficaces para su logro y una comprensión más completa de su impacto en la salud humana, merecen los mejores esfuerzos de los científicos del ejercicio.
Notas a pie de página
1 Utilizamos los términos cazador-recolector, recolector-cazador, y forrajeador de manera intercambiable
2 Hominoides: antepasados comunes de Homínidos y Póngidos.
3 Promedio en las tribus !Kung y Ache (104 kj/kg/d).
Referencias
1. Aiello LC, Wheeier PE (1995). The expensive-tissue hypothesis. Current Anthropol; 36: 199-221
2. American College of Sports Medicine (1995). ACSM Guidelines for Exercise Testing and Prescription. Kenney WL, editor. Baltimore: Williams &Wilkens. 153-67
3. Ástrand P-0. Whole body metabolism (1988). In: Horton E. Terjung R. editors. Exercise, Nutrition, and Energy Metabolism. New York:. MacMillan. 1-8
4. Baumann JE (1926). Observations on the strength of the chimpanzee and its implications. Mammol 7: 1 -9
5. Behrensmeyer AK, Cooke HBS (1985). Paleoenvironments, stratigraphy, and taphonomy in the African Pliocene and early Pleistocene. In: Delson E. editor. Ancestors: The Hard Evidence. New York: LissAlanR. 60-2
6. Blair SN (1993). C.H. McCloy Lecture: Physical activity, physical fitness, and health. Res Quart Exerc Sport; 64: 365 - 76
7. Blair SN (1995). Exercise prescription for health. Quest; 47; 333^ 53
8. Blair SN, Kohl H, Gordon N. Paffenbarger R (1992). How much physical activity is good for health?. Am Rev Public Health; 13: 99-126
9. Brooks GA, Fahey TD, White TP (1996). Exercise Physiology: Human Bioenergetics and Its Applications. Mountain View. CA: May-field Publishing Company: 517-8
10. Brown F, Harris J, Leakey R, Walker A (1985). Early Homo erectas skeleton from west lake Turkana. Kenya. Nature; 316: 788-92
11. Carrier DR (1984). The energetic paradox of human running and homi-nid evolution. Current Anthropol; 25: 483-95
12. Cavagna GA, Hegtund NC. Taylor CR (1997). Mechanical work in terrestrial locomotion: two basic mechanisms for minimizing energy expenditure. Am j Physiol; 233: R243 -R61
13. Cavagna GA (1976). Thys H, Zamboni A.The sources of external work in level walking and running. J Physiol; 262: 639-57
14. Chaplin G (1994). Jablonski NG. Cable NT. Physiology, thermoregula-tion and bipedalism. J. Hum Evolut; 27: 497-510
15. De Menocal PB (1995). Plio-Pleistocene African climate. Science; 270:53-9
16. DeQuincey T (1980). In: Davies H, editor. William Wordsworth. A Biography. London: Weidenfeld and Nicolson: 203
17. Eaton SB, Shostak M, Konner M (1988). The First Fitness Formula. In: The Paleolithic Prescription. New York: HarperSj Row: 168-99
18. Eaton SB, Konner M, Shostak M (1988). Stone agers in the fast lane: chronic degenerative diseases in evolutionary perspective. Am JMed; 84: 739-49
19. Finch G (1943). The bodily strength of chimpanzees. J Mammal; 24:224-8
20. Foley R (1984). Early man and the red queen: tropical African community evolution and hominid adaptation. In: Foley R, editor. Hominid Evolution and Community Ecology. New York: Academic Press: 85 -110
21. Foley R (1987). Another Unique Species: Patterns in Human Evolutionary Ecology. Harlow: Longman
22. Foley R, Lee PC (1989). Finite social space, evolutionary pathways, and reconstructing hominid behavior. Science; 243: 901-6
23. Gowlett JJ (1984). Mental abilities of early man: a look at some hard evidence. In: Foley R. editor. Hominid Evolution and Community Ecology. New York: Academic Press; 167-92
24. Helmirick SP (1991). Ragland D, Leung R. Physical activity and reduced occurrence of non-insulin-dependent diabetes mellitus. N Engl JMed; 325: 147-52
25. Heyward VH (1991). Advanced Fitness Assessment & Exercise Prescription. Champaign, !L: Human Kinetic Publishers: 326-31
26. Johanson DC (1996). Face-to-face with Lucy´s family. Nat Geogr; 189(3): 96-117
27. Kleiber M (1961). The Fire of Life. An Introduction to Animai Energetics. New York: Wiley: 212
28. Leakey MD (1971). Oiduvai Gorge, Vol 3. Excavations in Beds I and II, 1960-1963. Cambridge: Cambridge University Press
29. Leakey MD, Feibel CS (1995). McDougall I, Walker A. New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya. Nature; 376: 565-71
30. Lee RB (1979). The IKung San. Men, Women, and Work in a Foraging Society. London: Cambridge University Press: 313-7
31. Leon AS, Norstrom j (1995). Evidence of the role of physica! activity and cardiorespiratory fitness in the prevention of coronary heart disease. Quest; 47: 311-9
32. Leonard WR (1992). Robertson ML Nutritional requirements and human evolution: a bioenergetics model. Am J Hum Biol; 4: 179-95
33. Leonard WR, Robertson ML (1994). Evolutionary perspectives on human nutrition: the influence of brain and body size on diet and metabolism. Am j Hum Biol; 6: 77 - 88
34. Leslie P, Bindon J, Baker P (1984). Caloric requirement of human populations: a model. Human Ecology; 12: 137-62
35. Lovejoy CO (1988). Evolution of human walking. Sci Am: 259: 118-25
36. McArdle WD, Katch FI, Katch VL Exercise Physiology (1991). Energy, Nutrition and Human Performance. Philadelphia: Lea 8j Febiger: 421-51
37. Milton K (1993). Diet and primate evolution. Sci Am: 269: 86-93
38. Morris JN, Clayton DC, Everitt MG, Semmence AM (1990). Burgess EH. Exercise in leisure-time: coronary attack and death rate. Br Heart j: 63: 325-34
39. National Research Council (1989). Diet and Health. Implications for Reducing Chronic Disease Risk. Washington DC. National Academy Press; 142:52
40. Newman RW (1970). Why is man such a sweaty, thirsty, naked animal?. A speculative review: Hum Biol; 42:12-27
41. Paffenbarger RS (1981). Wing AL, Hyde RT. Chronic disease in former college students. XVI. Physical activity as an index of heart attack risk in college alumni. AmJ Epidemiol; 108: 161-75
42. Paffenbarger RS, Hyde R. Wing A, Hsieh CC (1986). Physical activity, all-cause mortality and longevity of college alumni. N Engl J Med:314:605-13
43. Rode A, Shephard RJ (1994). Predictions of body fat content ¡n an Inuit community. Am J Hum Biol: 6: 249 - 54
44. Rode A, Shephard RJ (1994). Physiological consequences of acculturation: a 20-year study of fitness in an Inuit community. Eur J Appl Physiol: 69: 516-24
45. Rodman PS, McHenry HM (1980). Bioenergetics and the origin of hominid bipedalism. Am j Phys Anthropol; 52: 103-6
46. Ruff CB (1991). Climate and body shape in hominid evolution. J Hum Evolut: 21: 81-105
47. Ruff CB.Trinkaus E, Walker A, Larsen'CS (1993). Postcranial robusticity in Homo: I: temporal trends and mechanical interpretation. Am j Phys Anthropol: 91: 21 - 53
48. Sahlins MD (1968). Notes on the original affluent society. In: Lee RB, DeVore 1. editors. Man the Hunter. Chicago: Aldine: 85-9
49. Shephard RJ (1978). Human physiological work capacity. Cambrigde: Cambridge University Press: 111: 56-65
50. Shipman P (1986). Scavenging or hunting in early hominids: theoretical framework and tests. Am Anthropol: 88: 27-43
51. Shvartz E, Reibold RC (1990). Aerobic fitness norms for males and females aged 6 to 75 years: a review. Aviat Space Environ Med: 61:3-11
52. Slattery ML, Jacobs DR, Nichaman MZ (1989). Leisure-time physical activity and coronary heart disease death: the US railroad study. Circulation: 79: 304-11
53. Stern JT, Susman RL (1983). The locomotor anatomy of Australopithecus afarensis. AmJ Phys Anthropol: 60: 279-317
54. Physical Activity (1998). Energy Expenditure and Fitness: An Evolutionary Perspective. Int. j. Sports Med. 19
55. Stringer CB (1984). The origin of anatomically modern humáns in western Europe, ¡n: Smith FH, Spencer F, editors. The Origin of Modern Humans: a World Survey of the Fossil Evidence. New York: Liss, Alan: 51-135'
56. Taylor CR, Heglund NC, Maloiy GMO (1982). Energetics and mechanics of terrestrial locomotion. l.J Exp Biol; 97: 1 -21
57. Taylor CR, Rowntree Vj (1973). Running on two or on four legs: which consumes more energy? . Science: 179:186-7
58. Tucker VA (1975). The cost of moving about. Am Scientist; 63: 413-9
59. Vigilant L, Stoneking M, Harpending H, Hawkes K, Wilson AC (1991). African populations and the evolution of human mitochondria! . DNA.Science; 253: 1503-7
60. Wheeler PE (1991). The influence of bipedalism on the energy and water budgets of early hominids. J Hum Evolut: 21:117-36
61. Wheeler PE (1992). The influence of the loss of functional body hair on the water budgets of early hominids. J Hum Evolut: 23: 379-88
62. Wheeier PE (1993). The influence of stature and body form on hominid energy and water budgets; a comparison of Australopithecus and early Homo physiques. J Hum Evolut; 24:13-28
63. Wilson AC, Cann RL (1992). The recent African genesis of humans. Sci Am; 266: 68-73
Cita Original
L Cordain, R. W. Gotshall, S. Boyd Eaton, S. Boyd Eaton III. Physical Activity, Energy Expenditure and Fitness: An Evolutionary Perspective. Int.J. Sports Med., Vol. 19, pp.328-335,1998
Cita en PubliCE
R. W Gotshall, L Cordain, Boyd Eaton Boyd Eaton III (1998). Actividad Física, Gasto de Energía y Aptitud Física: Una Perspectiva Evolutiva . .https://g-se.com/actividad-fisica-gasto-de-energia-y-aptitud-fisica-una-perspectiva-evolutiva--1416-sa-R57cfb2720af5f